

The genome of the clonal raider ant Cerapachys biroi
- PMID: 24508170
- PMCID: PMC3961065
- DOI: 10.1016/j.cub.2014.01.018
The genome of the clonal raider ant Cerapachys biroi
Abstract
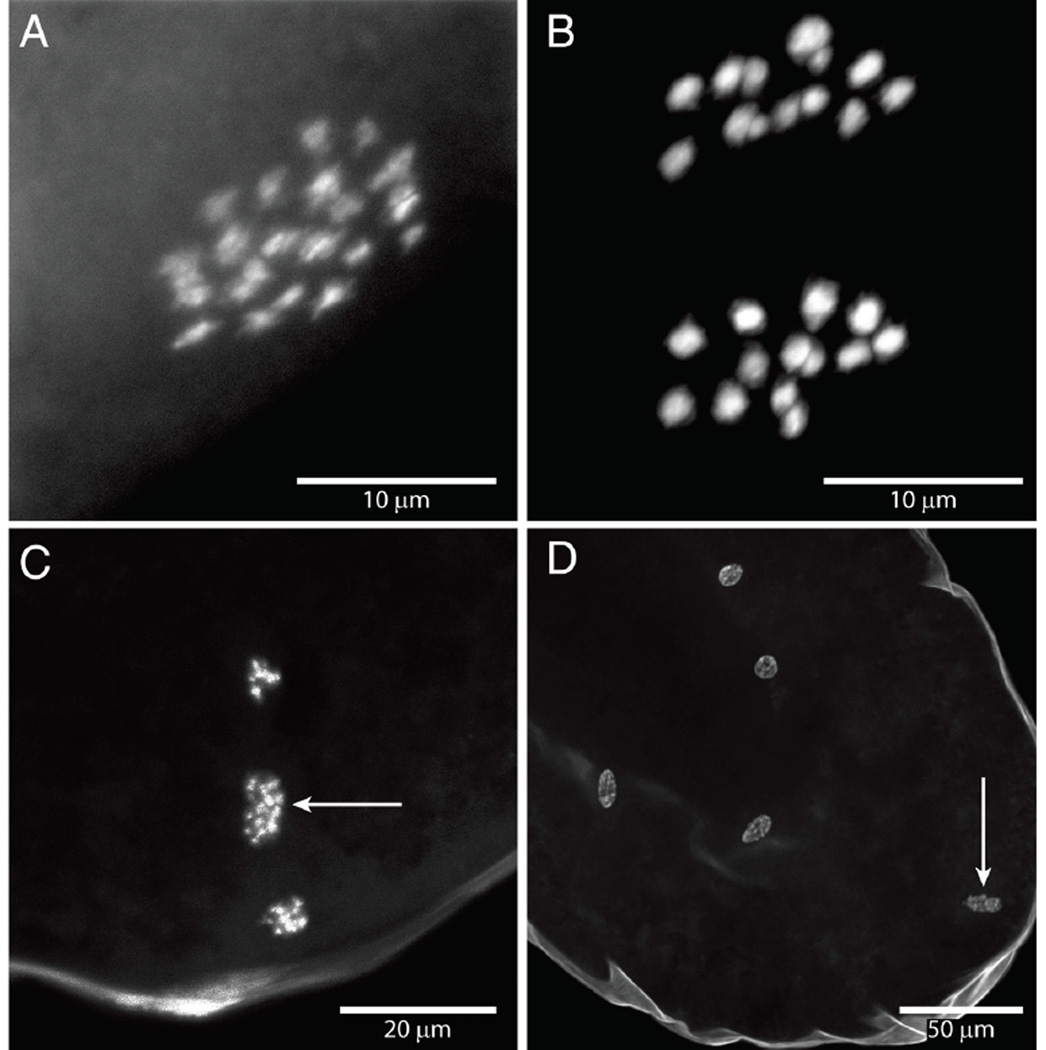
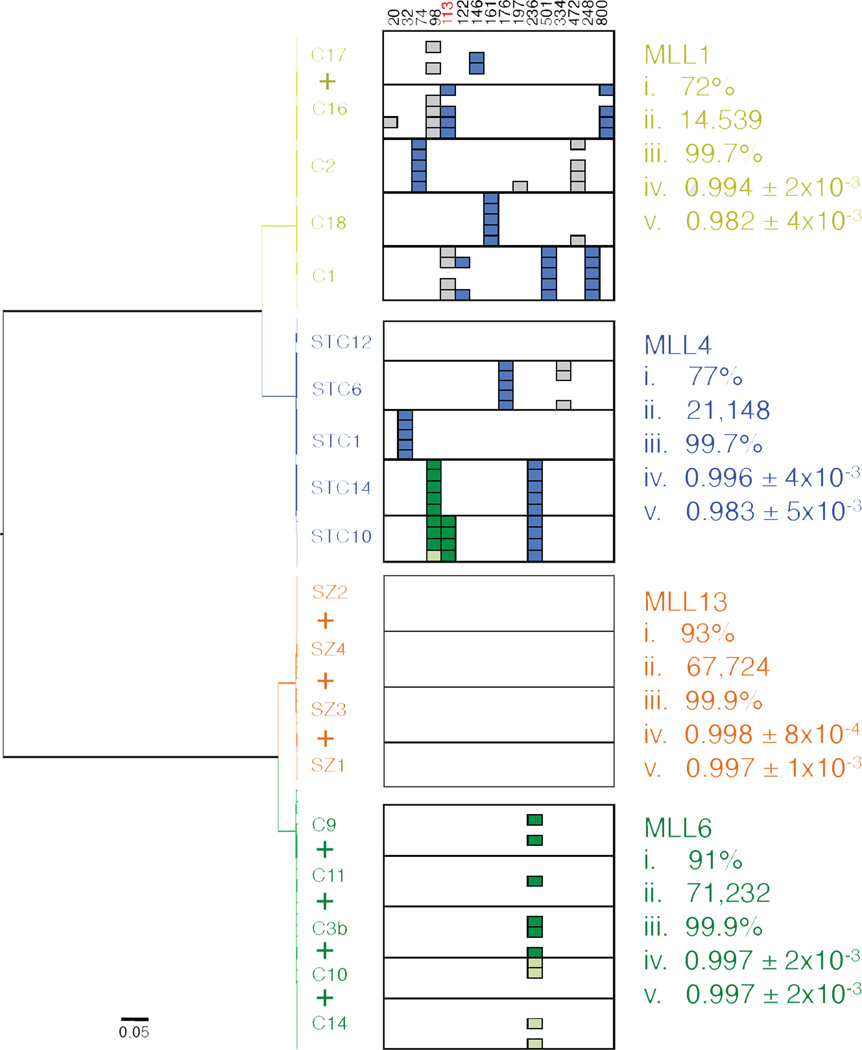
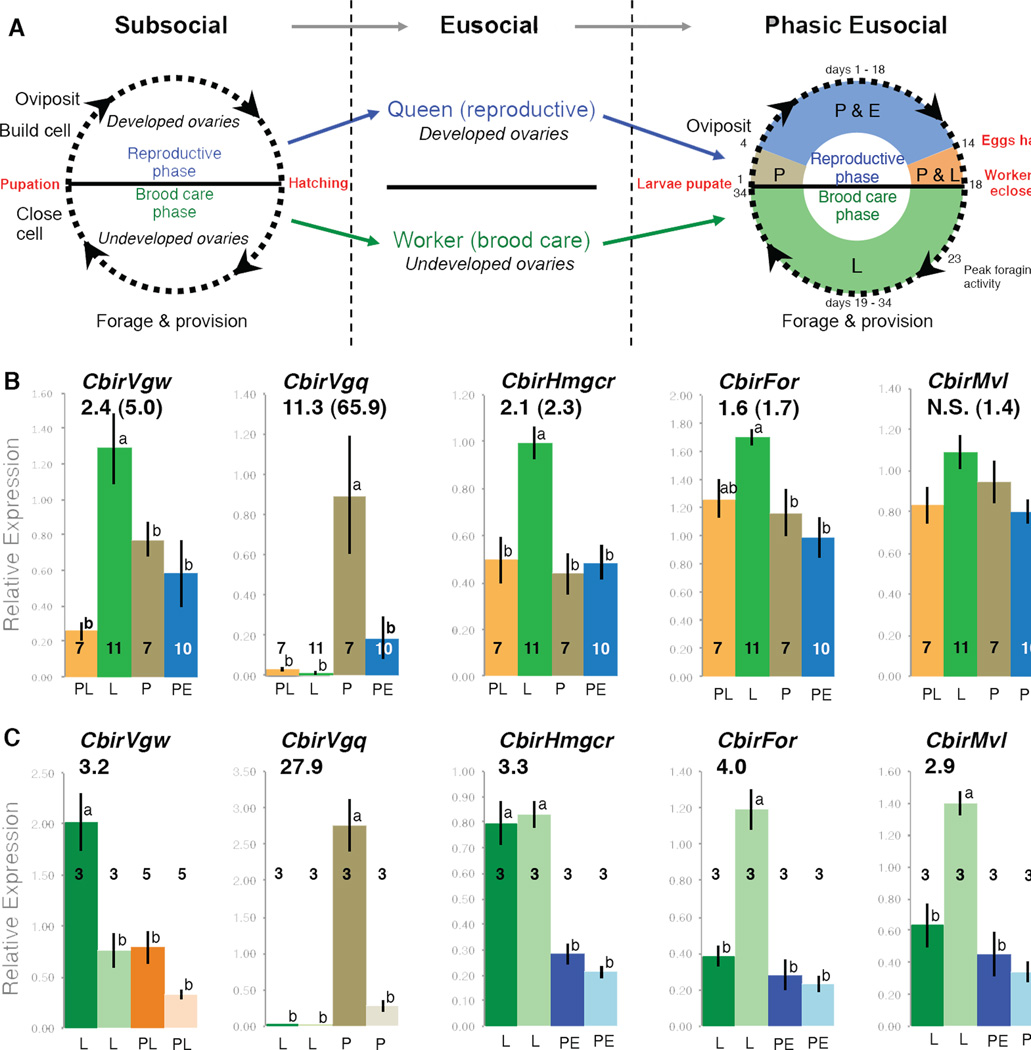
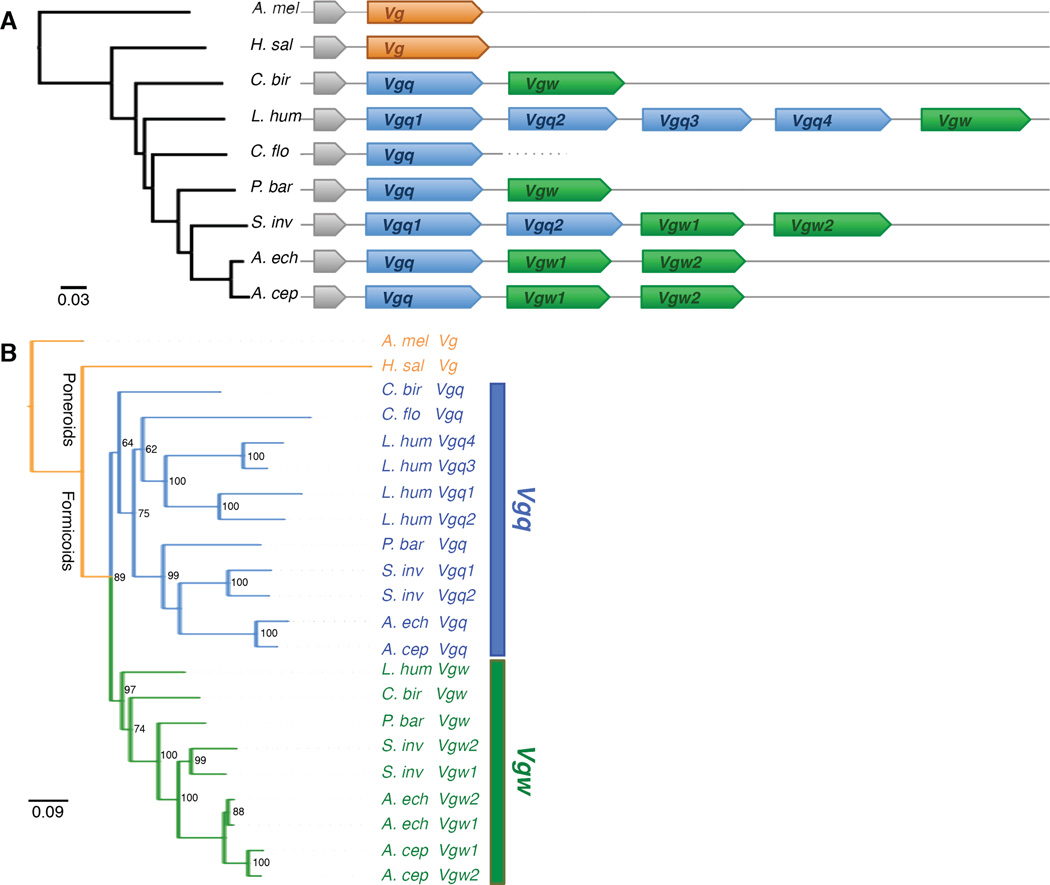
Social insects are important models for social evolution and behavior. However, in many species, experimental control over important factors that regulate division of labor, such as genotype and age, is limited. Furthermore, most species have fixed queen and worker castes, making it difficult to establish causality between the molecular mechanisms that underlie reproductive division of labor, the hallmark of insect societies. Here we present the genome of the queenless clonal raider ant Cerapachys biroi, a powerful new study system that does not suffer from these constraints. Using cytology and RAD-seq, we show that C. biroi reproduces via automixis with central fusion and that heterozygosity is lost extremely slowly. As a consequence, nestmates are almost clonally related (r = 0.996). Workers in C. biroi colonies synchronously alternate between reproduction and brood care, and young workers eclose in synchronized cohorts. We show that genes associated with division of labor in other social insects are conserved in C. biroi and dynamically regulated during the colony cycle. With unparalleled experimental control over an individual's genotype and age, and the ability to induce reproduction and brood care, C. biroi has great potential to illuminate the molecular regulation of division of labor.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures

Similar articles
-
Robust DNA Methylation in the Clonal Raider Ant Brain.Curr Biol. 2016 Feb 8;26(3):391-5. doi: 10.1016/j.cub.2015.12.040. Epub 2016 Jan 21. Curr Biol. 2016. PMID: 26804553 Free PMC article.
-
Clonal raider ant brain transcriptomics identifies candidate molecular mechanisms for reproductive division of labor.BMC Biol. 2018 Aug 13;16(1):89. doi: 10.1186/s12915-018-0558-8. BMC Biol. 2018. PMID: 30103762 Free PMC article.
-
Genetic distance and age affect the cuticular chemical profiles of the clonal ant Cerapachys biroi.J Chem Ecol. 2014 May;40(5):429-38. doi: 10.1007/s10886-014-0428-y. Epub 2014 Apr 23. J Chem Ecol. 2014. PMID: 24756691
-
The genomic impact of 100 million years of social evolution in seven ant species.Trends Genet. 2012 Jan;28(1):14-21. doi: 10.1016/j.tig.2011.08.005. Epub 2011 Oct 5. Trends Genet. 2012. PMID: 21982512 Free PMC article. Review.
-
Genes and genomic processes underpinning the social lives of ants.Curr Opin Insect Sci. 2018 Feb;25:83-90. doi: 10.1016/j.cois.2017.12.001. Epub 2017 Dec 20. Curr Opin Insect Sci. 2018. PMID: 29602366 Review.
Cited by
-
Genomic Signals of Adaptation towards Mutualism and Sociality in Two Ambrosia Beetle Complexes.Life (Basel). 2018 Dec 22;9(1):2. doi: 10.3390/life9010002. Life (Basel). 2018. PMID: 30583535 Free PMC article.
-
Evolution of five environmentally responsive gene families in a pine-feeding sawfly, Neodiprion lecontei (Hymenoptera: Diprionidae).Ecol Evol. 2023 Oct 1;13(10):e10506. doi: 10.1002/ece3.10506. eCollection 2023 Oct. Ecol Evol. 2023. PMID: 37791292 Free PMC article.
-
Target enrichment of ultraconserved elements from arthropods provides a genomic perspective on relationships among Hymenoptera.Mol Ecol Resour. 2015 May;15(3):489-501. doi: 10.1111/1755-0998.12328. Epub 2014 Sep 29. Mol Ecol Resour. 2015. PMID: 25207863 Free PMC article.
-
Stress and early experience underlie dominance status and division of labour in a clonal insect.Proc Biol Sci. 2018 Aug 29;285(1885):20181468. doi: 10.1098/rspb.2018.1468. Proc Biol Sci. 2018. PMID: 30158313 Free PMC article.
-
Evolution, developmental expression and function of odorant receptors in insects.J Exp Biol. 2020 Feb 7;223(Pt Suppl 1):jeb208215. doi: 10.1242/jeb.208215. J Exp Biol. 2020. PMID: 32034042 Free PMC article. Review.
References
-
- Sokolowski MB. Social interactions in “simple” model systems. Neuron. 2010;65:780–794. - PubMed
-
- LeBoeuf AC, Benton R, Keller L. The molecular basis of social behavior: models, methods and advances. Curr. Opin. Neurobiol. 2013;23:3–10. - PubMed
-
- Smith CR, Toth AL, Suarez AV, Robinson GE. Genetic and genomic analyses of the division of labour in insect societies. Nat. Rev. Genet. 2008;9:735–748. - PubMed
-
- Teseo S, Kronauer DJC, Jaisson P, Châline N. Enforcement of reproductive synchrony via policing in a clonal ant. Curr. Biol. 2013;23:328–332. - PubMed
-
- Ravary F, Jahyny B, Jaisson P. Brood stimulation controls the phasic reproductive cycle of the parthenogenetic ant Cerapachys biroi. Insectes Soc. 2006;53:20–26.
Publication types
MeSH terms
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
