

CXCL5-secreting pulmonary epithelial cells drive destructive neutrophilic inflammation in tuberculosis
- PMID: 24509076
- PMCID: PMC3934185
- DOI: 10.1172/JCI72030
CXCL5-secreting pulmonary epithelial cells drive destructive neutrophilic inflammation in tuberculosis
Abstract
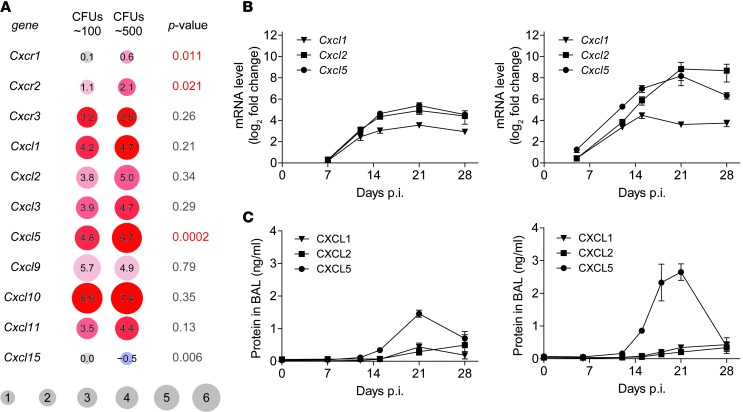
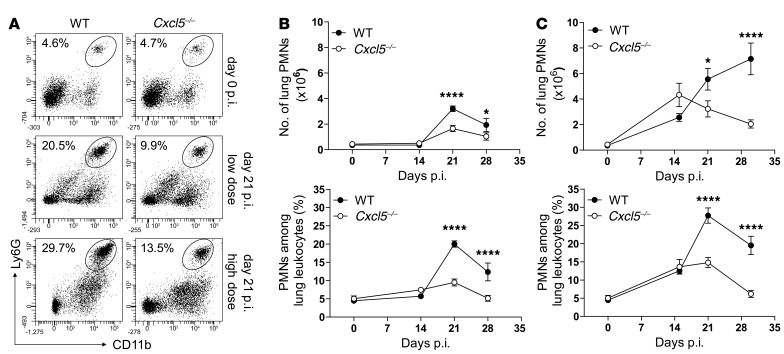
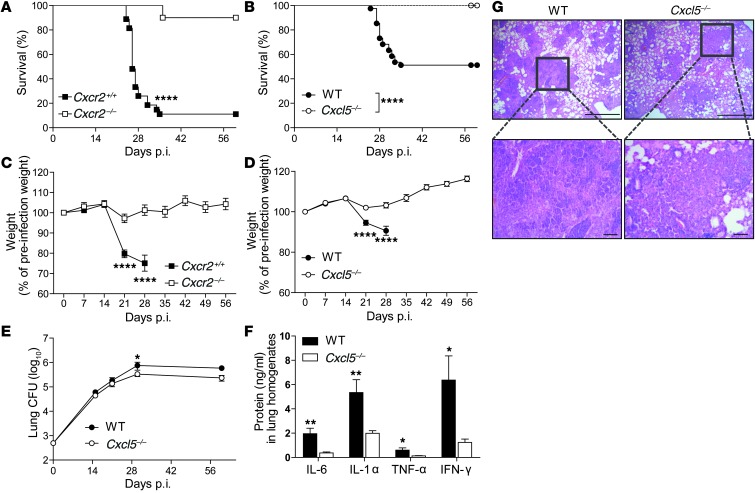
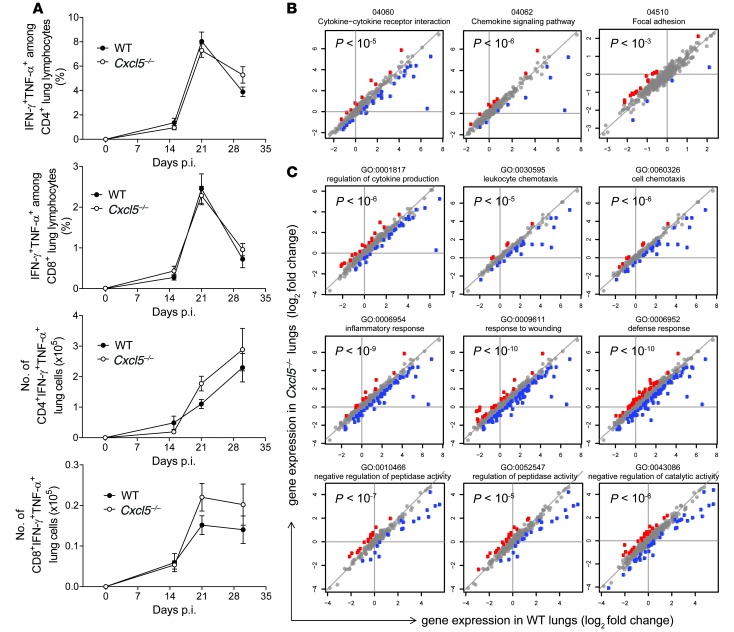
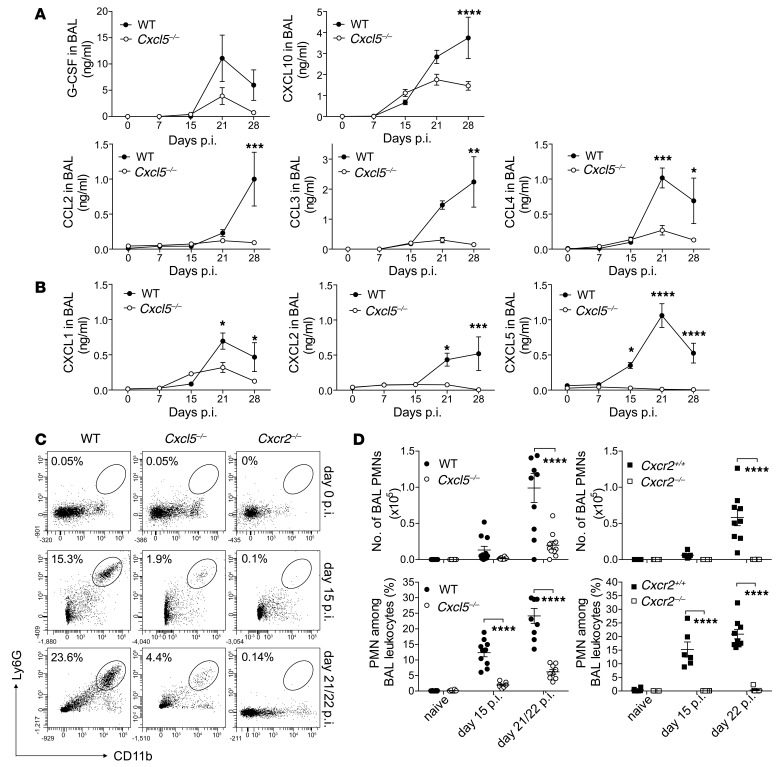
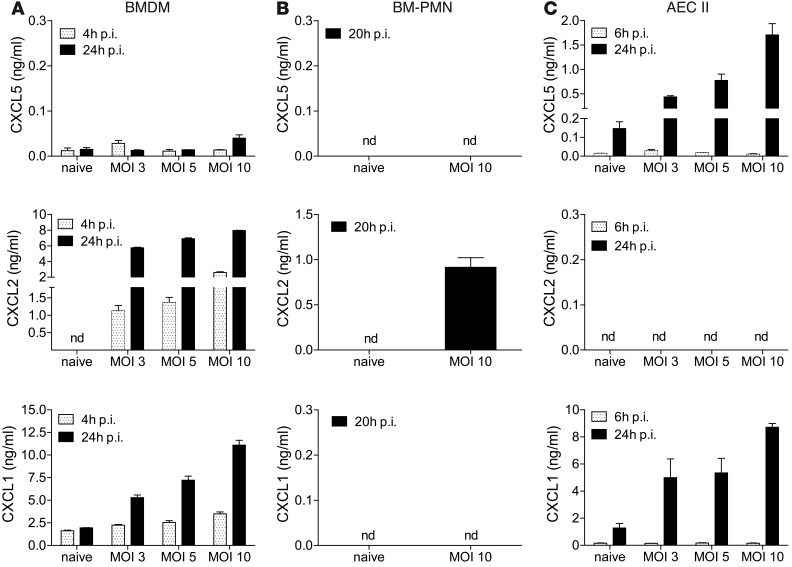
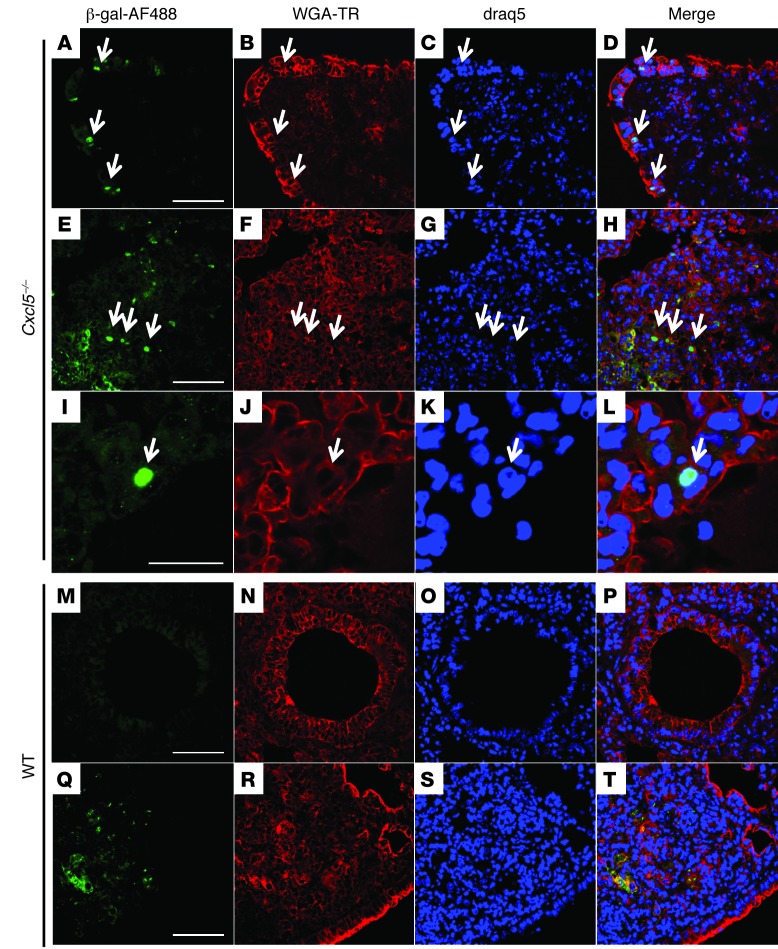
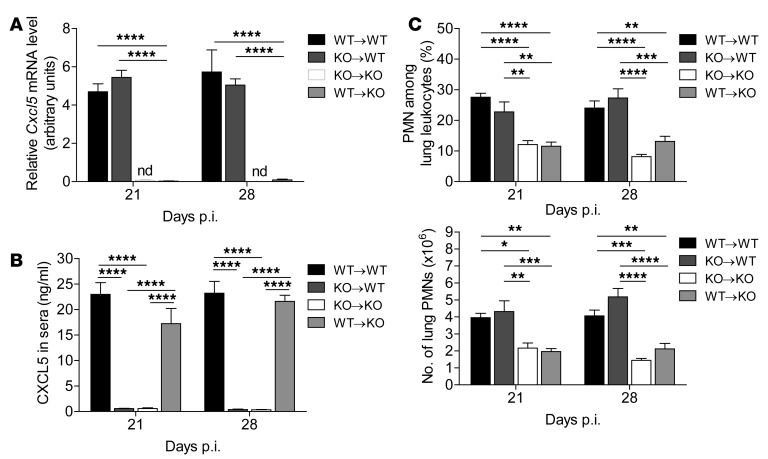
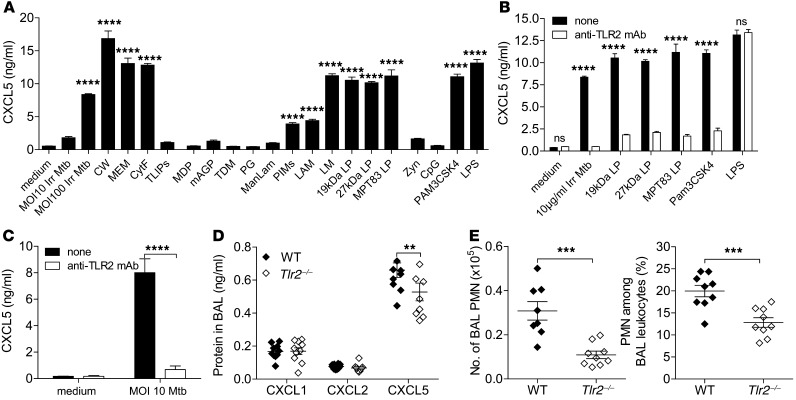
Successful host defense against numerous pulmonary infections depends on bacterial clearance by polymorphonuclear leukocytes (PMNs); however, excessive PMN accumulation can result in life-threatening lung injury. Local expression of CXC chemokines is critical for PMN recruitment. The impact of chemokine-dependent PMN recruitment during pulmonary Mycobacterium tuberculosis infection is not fully understood. Here, we analyzed expression of genes encoding CXC chemokines in M. tuberculosis-infected murine lung tissue and found that M. tuberculosis infection promotes upregulation of Cxcr2 and its ligand Cxcl5. To determine the contribution of CXCL5 in pulmonary PMN recruitment, we generated Cxcl5(-/-) mice and analyzed their immune response against M. tuberculosis. Both Cxcr2(-/-) mice and Cxcl5(-/-) mice, which are deficient for only one of numerous CXCR2 ligands, exhibited enhanced survival compared with that of WT mice following high-dose M. tuberculosis infection. The resistance of Cxcl5(-/-) mice to M. tuberculosis infection was not due to heightened M. tuberculosis clearance but was the result of impaired PMN recruitment, which reduced pulmonary inflammation. Lung epithelial cells were the main source of CXCL5 upon M. tuberculosis infection, and secretion of CXCL5 was reduced by blocking TLR2 signaling. Together, our data indicate that TLR2-induced epithelial-derived CXCL5 is critical for PMN-driven destructive inflammation in pulmonary tuberculosis.
Figures
Similar articles
-
Toll-like Receptor 2 Prevents Neutrophil-Driven Immunopathology during Infection with Mycobacterium tuberculosis by Curtailing CXCL5 Production.Infect Immun. 2019 Feb 21;87(3):e00760-18. doi: 10.1128/IAI.00760-18. Print 2019 Mar. Infect Immun. 2019. PMID: 30559223 Free PMC article.
-
A protective role for ELR+ chemokines during acute viral encephalomyelitis.PLoS Pathog. 2009 Nov;5(11):e1000648. doi: 10.1371/journal.ppat.1000648. Epub 2009 Nov 6. PLoS Pathog. 2009. PMID: 19893623 Free PMC article.
-
Phospholipase Cε plays a crucial role in neutrophilic inflammation accompanying acute lung injury through augmentation of CXC chemokine production from alveolar epithelial cells.Respir Res. 2019 Jan 11;20(1):9. doi: 10.1186/s12931-019-0975-4. Respir Res. 2019. PMID: 30634975 Free PMC article.
-
Perspectives on host adaptation in response to Mycobacterium tuberculosis: modulation of inflammation.Semin Immunol. 2014 Dec;26(6):533-42. doi: 10.1016/j.smim.2014.10.002. Epub 2014 Oct 25. Semin Immunol. 2014. PMID: 25453228 Review.
-
Neutrophils in tuberculosis--first line of defence or booster of disease and targets for host-directed therapy?Pathog Dis. 2016 Apr;74(3):ftw012. doi: 10.1093/femspd/ftw012. Epub 2016 Feb 22. Pathog Dis. 2016. PMID: 26903072 Review.
Cited by
-
The Src-Family Kinases Hck and Fgr Regulate Early Lipopolysaccharide-Induced Myeloid Cell Recruitment into the Lung and Their Ability To Secrete Chemokines.J Immunol. 2015 Sep 1;195(5):2383-95. doi: 10.4049/jimmunol.1402011. Epub 2015 Jul 31. J Immunol. 2015. PMID: 26232427 Free PMC article.
-
Immunologic Role of Innate Lymphoid Cells against Mycobacterial tuberculosis Infection.Biomedicines. 2022 Nov 6;10(11):2828. doi: 10.3390/biomedicines10112828. Biomedicines. 2022. PMID: 36359348 Free PMC article. Review.
-
An expansion of rare lineage intestinal microbes characterizes rheumatoid arthritis.Genome Med. 2016 Apr 21;8(1):43. doi: 10.1186/s13073-016-0299-7. Genome Med. 2016. PMID: 27102666 Free PMC article.
-
Granulomas and Inflammation: Host-Directed Therapies for Tuberculosis.Front Immunol. 2016 Oct 24;7:434. doi: 10.3389/fimmu.2016.00434. eCollection 2016. Front Immunol. 2016. PMID: 27822210 Free PMC article. Review.
-
A siRNA targets and inhibits a broad range of SARS-CoV-2 infections including Delta variant.EMBO Mol Med. 2022 Apr 7;14(4):e15298. doi: 10.15252/emmm.202115298. Epub 2022 Feb 21. EMBO Mol Med. 2022. PMID: 35138028 Free PMC article.
References
-
- World Health Organization. Global Tuberculosis Report 2012. Geneva, Switzerland: World Health Organization; 2012.
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
