

Structural and mechanistic insight into Holliday-junction dissolution by topoisomerase IIIα and RMI1
- PMID: 24509834
- PMCID: PMC4292918
- DOI: 10.1038/nsmb.2775
Structural and mechanistic insight into Holliday-junction dissolution by topoisomerase IIIα and RMI1
Abstract
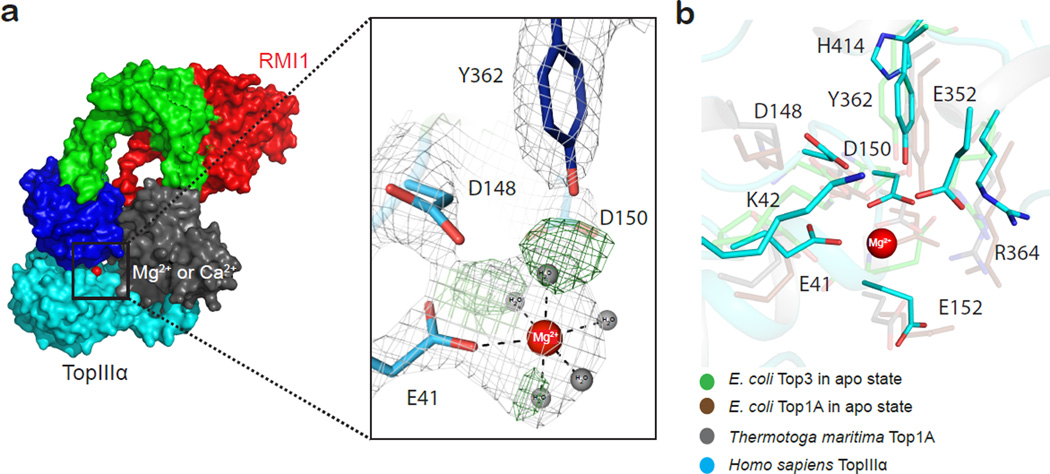
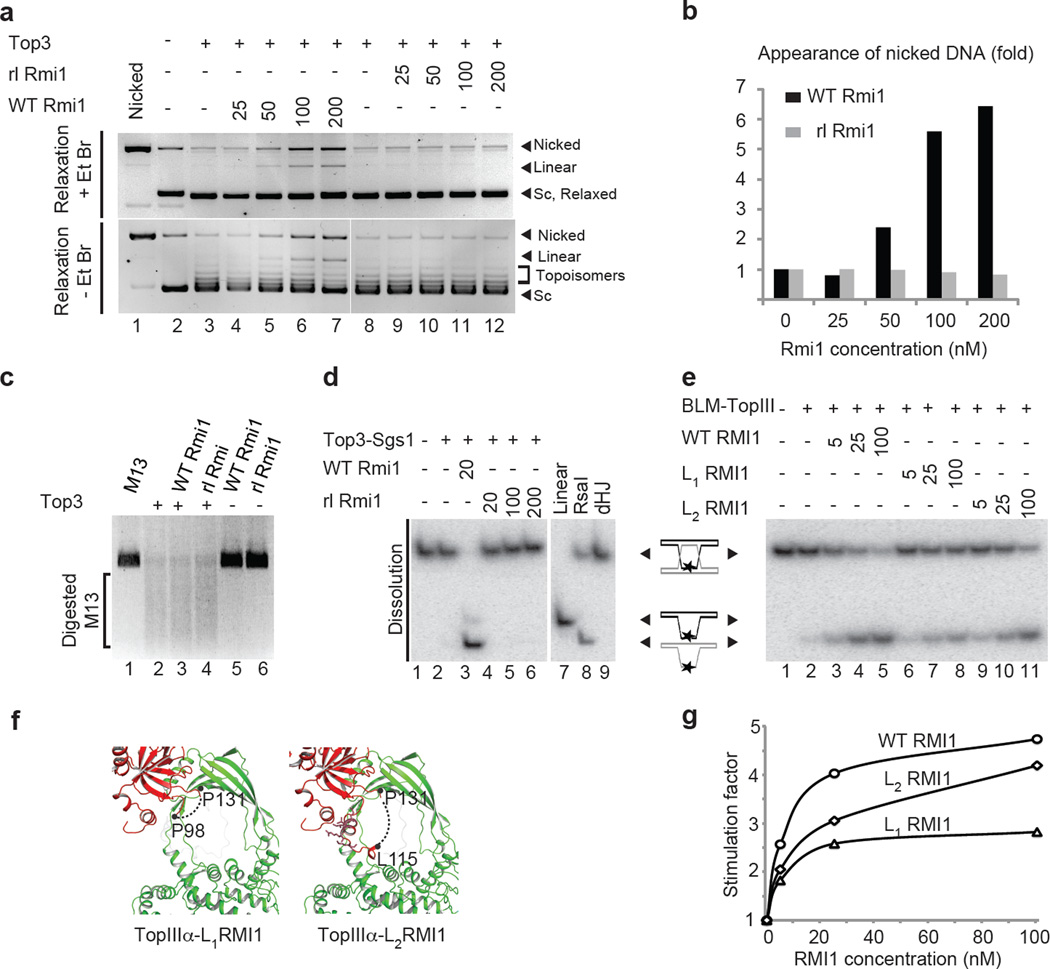
Repair of DNA double-strand breaks via homologous recombination can produce double Holliday junctions (dHJs) that require enzymatic separation. Topoisomerase IIIα (TopIIIα) together with RMI1 disentangles the final hemicatenane intermediate obtained once dHJs have converged. How binding of RMI1 to TopIIIα influences it to behave as a hemicatenane dissolvase, rather than as an enzyme that relaxes DNA topology, is unknown. Here, we present the crystal structure of human TopIIIα complexed to the first oligonucleotide-binding domain (OB fold) of RMI1. TopIII assumes a toroidal type 1A topoisomerase fold. RMI1 attaches to the edge of the gate in TopIIIα through which DNA passes. RMI1 projects a 23-residue loop into the TopIIIα gate, thereby influencing the dynamics of its opening and closing. Our results provide a mechanistic rationale for how RMI1 stabilizes TopIIIα-gate opening to enable dissolution and illustrate how binding partners modulate topoisomerase function.
Figures



References
References of the main text
-
- Petermann E, Helleday T. Pathways of mammalian replication fork restart. Nature reviews. Molecular cell biology. 2010;11:683–687. - PubMed
-
- Holliday R. A mechanism for gene conversion in fungi. Genetical research. 2007;89:285–307. - PubMed
-
- Szostak JW, Orr-Weaver TL, Rothstein RJ, Stahl FW. The double-strand-break repair model for recombination. Cell. 1983;33:25–35. - PubMed
-
- Heyer WD, Ehmsen KT, Solinger JA. Holliday junctions in the eukaryotic nucleus: resolution in sight? Trends in biochemical sciences. 2003;28:548–557. - PubMed
References for online methods
-
- Kabsch W. Automatic Processing of Rotation Diffraction Data from Crystals of Initially Unknown Symmetry and Cell Constants. Journal of Applied Crystallography. 1993;26:795–800.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
