

An evolutionarily conserved mechanism of calcium-dependent neurotoxicity in a zebrafish model of fetal alcohol spectrum disorders
- PMID: 24512079
- PMCID: PMC3999225
- DOI: 10.1111/acer.12360
An evolutionarily conserved mechanism of calcium-dependent neurotoxicity in a zebrafish model of fetal alcohol spectrum disorders
Abstract
Background: Fetal alcohol spectrum disorders (FASD) are a leading cause of neurodevelopmental disability. Nonhuman animal models offer novel insights into its underlying mechanisms. Although the developing zebrafish has great promise for FASD research, a significant challenge to its wider adoption is the paucity of clear, mechanistic parallels between its ethanol (EtOH) responses and those of nonpiscine, established models. Inconsistencies in the published pharmacodynamics for EtOH-exposed zebrafish, alongside the use of comparatively high EtOH doses, challenge the interpretation of this model's clinical relevance.
Methods: To address these limitations, we developed a binge, single-exposure model of EtOH exposure in the early zebrafish embryo.
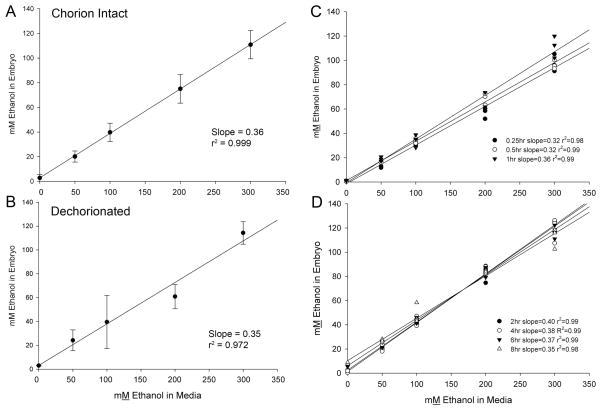
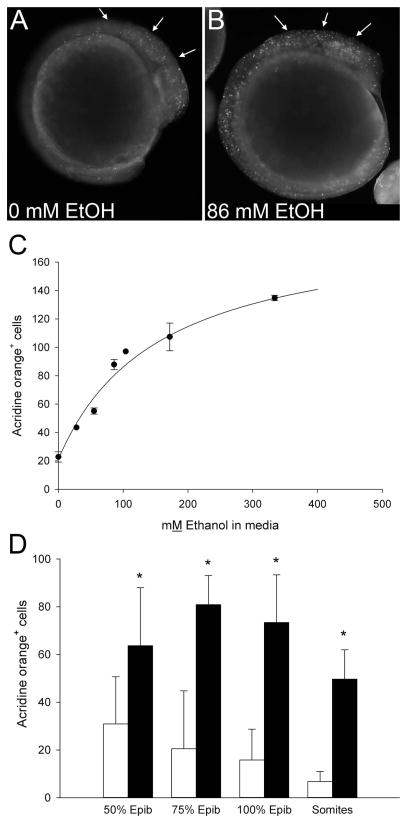
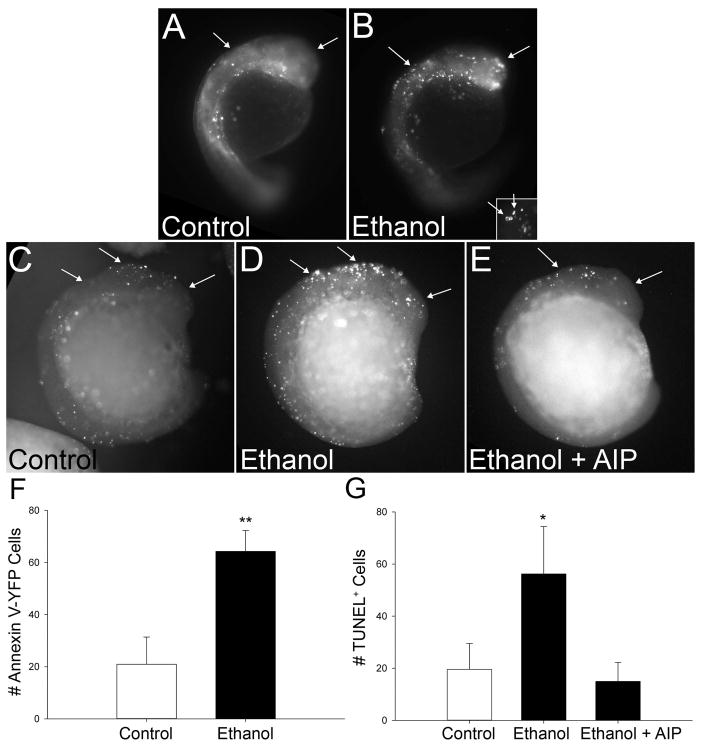
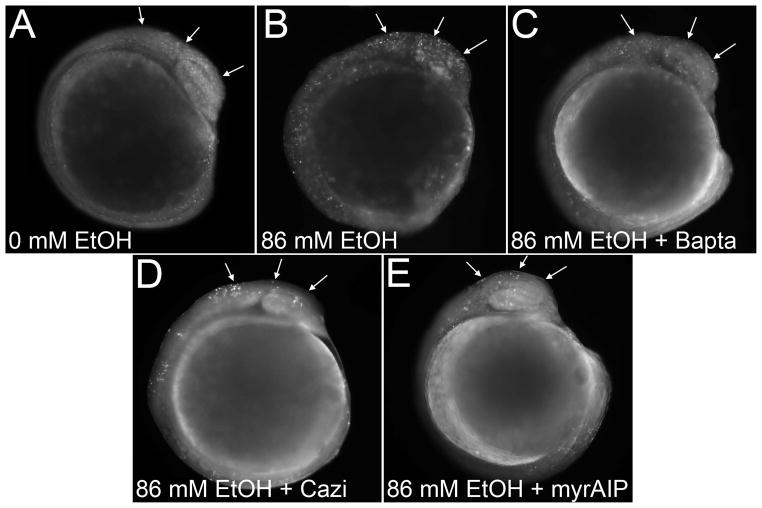
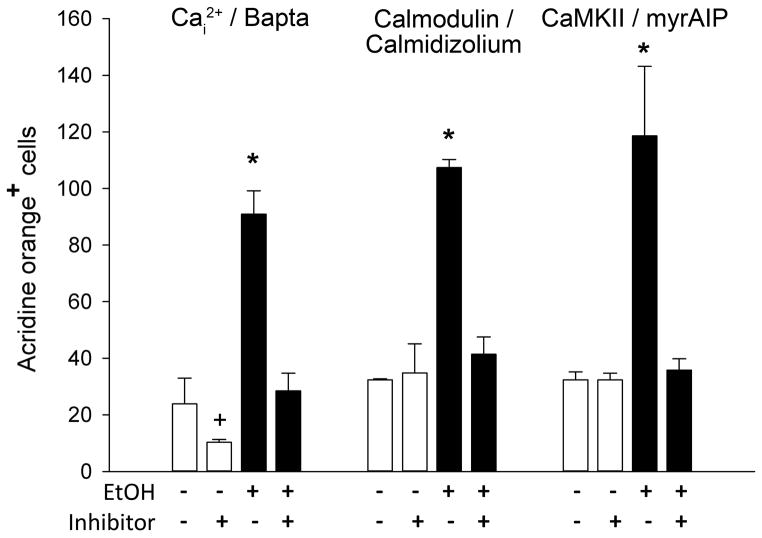
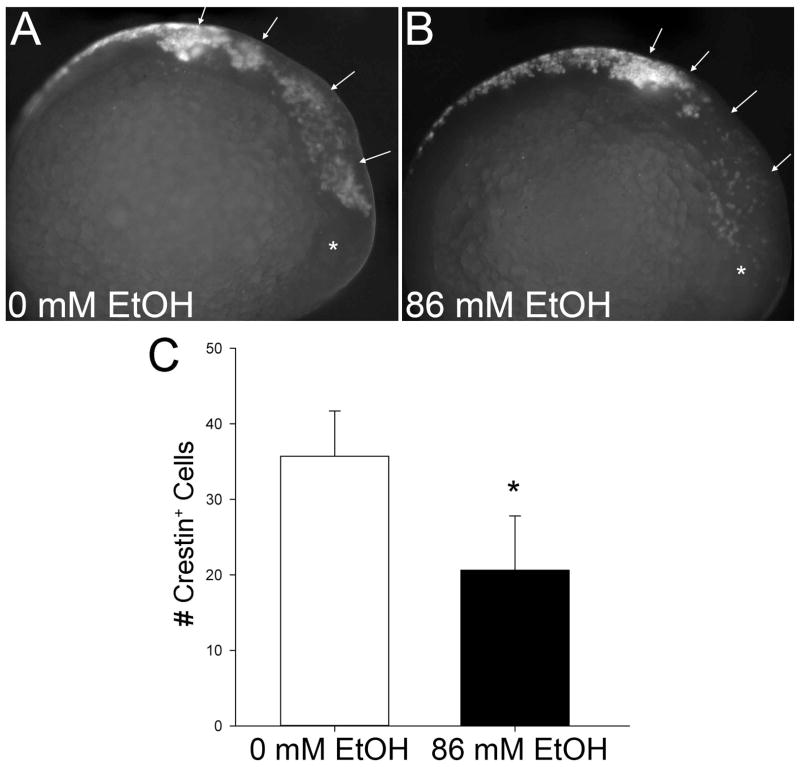
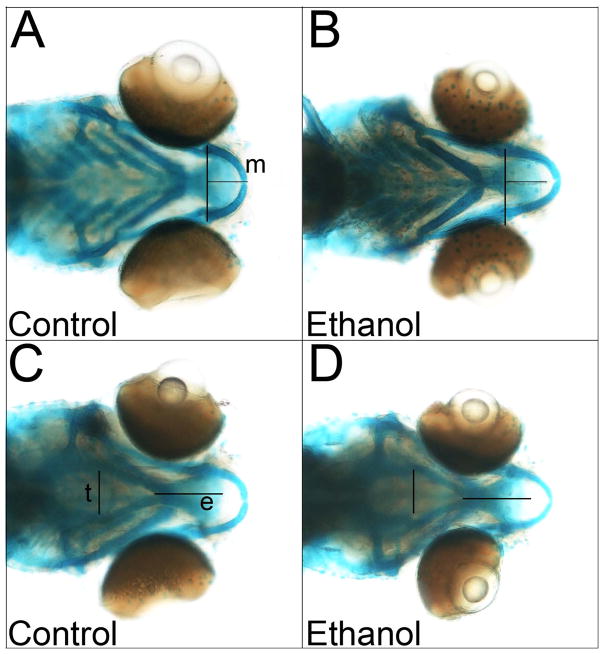
Results: Brief (3-hour) EtOH exposure is sufficient to cause significant neural crest losses and craniofacial alterations, with peak vulnerability during neurogenesis and early somitogenesis. These losses are apoptotic, documented using TUNEL assay and secA5-YFP-reporter fish. Apoptosis is dose dependent with an EC50 = 56.2 ± 14.3 mM EtOHint , a clinically relevant value within the range producing apoptosis in chick and mouse neural crest. This apoptosis requires the calcium-dependent activation of CaMKII and recapitulates the well-described EtOH signaling mechanism in avian neural crest. Importantly, we resolve the existing confusion regarding zebrafish EtOH kinetics. We show that steady-state EtOH concentrations within both chorion-intact and dechorionated embryos are maintained at 35.7 ± 2.8% of EtOHext levels across the range from 50 to 300 mM EtOHext , a value consistent with several published reports. Equilibrium is rapid and complete within 5 minutes of EtOH addition.
Conclusions: The calcium/CaMKII mechanism of EtOH's neurotoxicity is shared between an amniote (chick) and teleost fish, indicating that this mechanism is evolutionarily conserved. Our data suggest that EtOHext concentrations >2% (v/v) for chorion-intact embryos and 1.5% (v/v) for dechorionated embryos have limited clinical relevance. The strong parallels with established models endorse the zebrafish's relevance for mechanistic studies of EtOH's developmental neurotoxicity.
Keywords: Apoptosis; Calcium Signaling; Ethanol; Fetal Alcohol Spectrum Disorders; Neural Crest; Zebrafish.
Copyright © 2014 by the Research Society on Alcoholism.
Figures
Comment in
-
Commentary: catching a conserved mechanism of ethanol teratogenicity.Alcohol Clin Exp Res. 2014 Aug;38(8):2160-3. doi: 10.1111/acer.12484. Alcohol Clin Exp Res. 2014. PMID: 25156611 Free PMC article.
References
-
- Battaglia FC. Fetal alcohol syndrome diagnosis, epidimiology, prevention and treatment. Washington, D.C: National Academy Press; 1996.
-
- Bilotta J, Barnett JA, Hancock L, Saszik S. Ethanol exposure alters zebrafish development: a novel model of fetal alcohol syndrome. Neurotox Teratol. 2004;26:737–743. - PubMed
-
- Blader P, Strahle U. Ethanol impairs migration of the prechordal plate in the zebrafish embryo. Dev Biol. 1998;201:185–201. - PubMed
-
- Bradfield JY, West JR, Maider SE. Uptake and elimination of ethanol by young zebrafish embryos. Neurotox Teratol. 2006;28:629–633. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
