

An overview of mechanisms of redox signaling
- PMID: 24512843
- PMCID: PMC4048798
- DOI: 10.1016/j.yjmcc.2014.01.018
An overview of mechanisms of redox signaling
Abstract
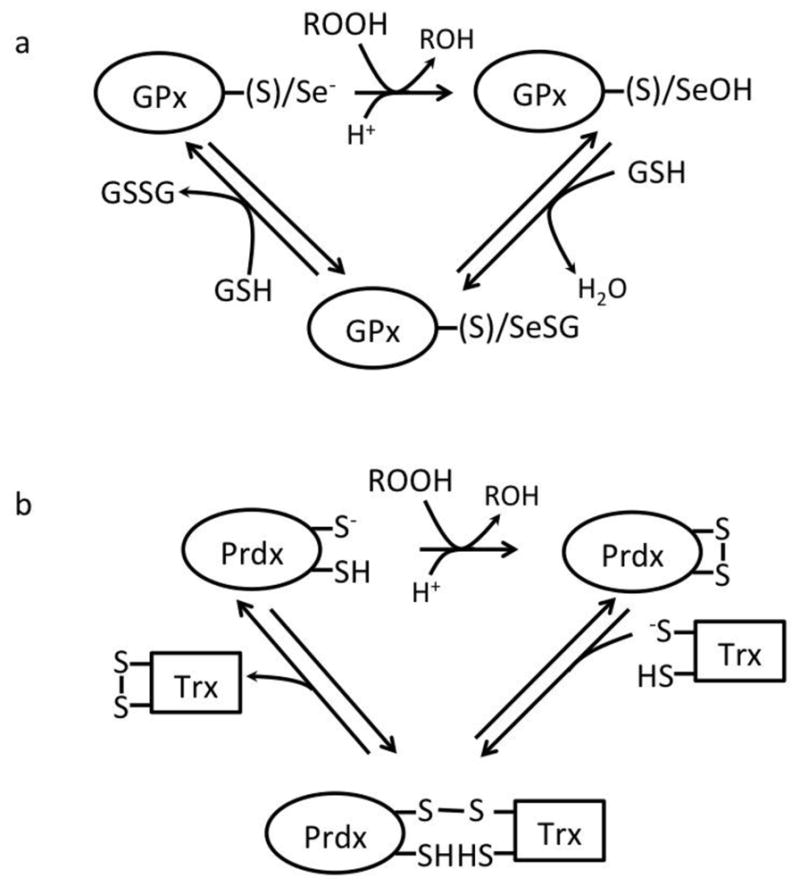
A principal characteristic of redox signaling is that it involves an oxidation-reduction reaction or covalent adduct formation between the sensor signaling protein and second messenger. Non-redox signaling may involve alteration of the second messenger as in hydrolysis of GTP by G proteins, modification of the signaling protein as in farnesylation, or simple non-covalent binding of an agonist or second messenger. The chemistry of redox signaling is reviewed here. Specifically we have described how among the so-called reactive oxygen species, only hydroperoxides clearly fit the role of a second messenger. Consideration of reaction kinetics and cellular location strongly suggests that for hydroperoxides, particular protein cysteines are the targets and that the requirements for redox signaling is that these cysteines are in microenvironments in which the cysteine is ionized to the thiolate, and a proton can be donated to form a leaving group. The chemistry described here is the same as occurs in the cysteine and selenocysteine peroxidases that are generally considered the primary defense against oxidative stress. But, these same enzymes can also act as the sensors and transducer for signaling. Conditions that would allow specific signaling by peroxynitrite and superoxide are also defined. Signaling by other electrophiles, which includes lipid peroxidation products, quinones formed from polyphenols and other metabolites also involves reaction with specific protein thiolates. Again, kinetics and location are the primary determinants that provide specificity required for physiological signaling although enzymatic catalysis is not likely involved. This article is part of a Special Issue entitled "Redox Signalling in the Cardiovascular System".
Keywords: 4-Hydroxynonenal; Hydrogen peroxide; Hydroperoxide; Peroxidases; Reactive oxygen species; Signal transduction.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures
References
-
- Toppo S, Flohé L, Ursini F, Vanin S, Maiorino M. Catalytic mechanisms and specificities of glutathione peroxidases: Variations of a basic scheme. Biochimica et biophysica acta. 2009;1790:1486–500. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
