

A fate-map for cranial sensory ganglia in the sea lamprey
- PMID: 24513489
- PMCID: PMC3928997
- DOI: 10.1016/j.ydbio.2013.10.021
A fate-map for cranial sensory ganglia in the sea lamprey
Abstract
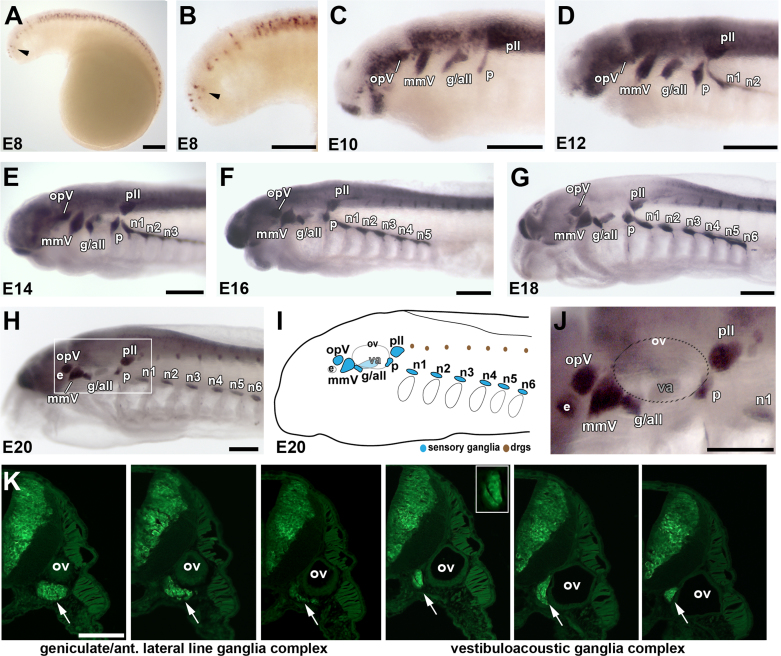
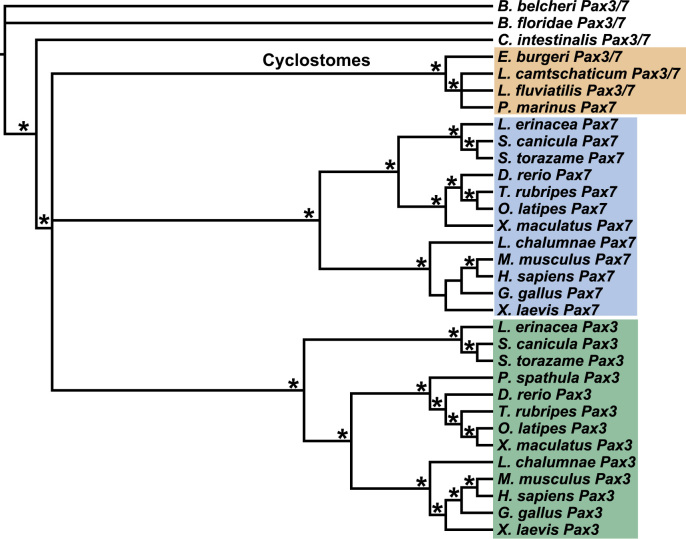
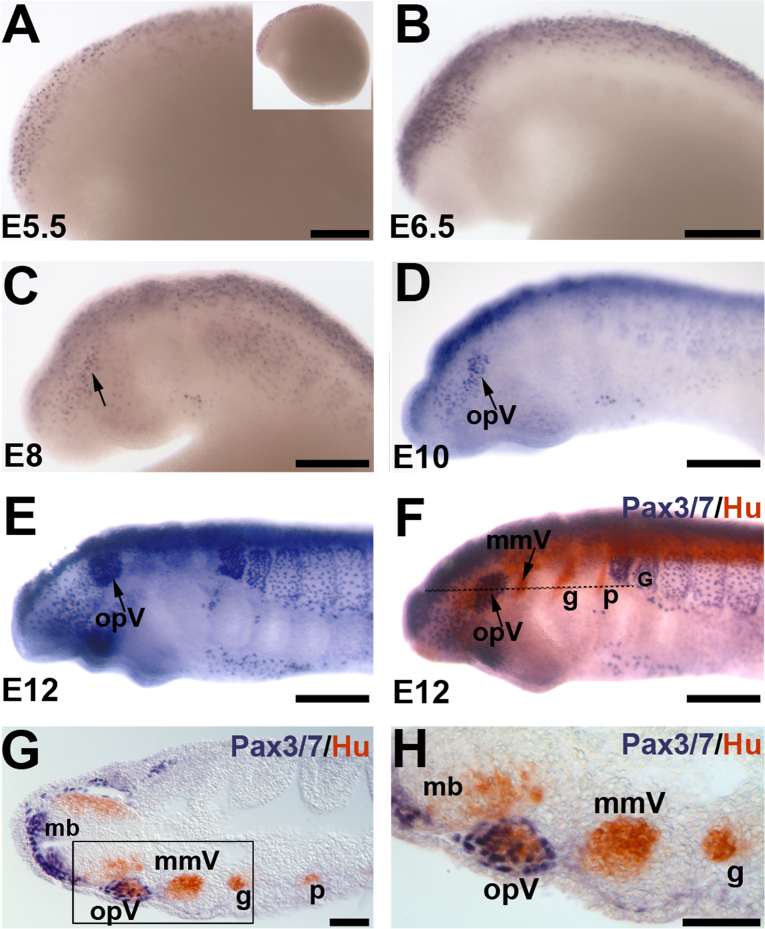
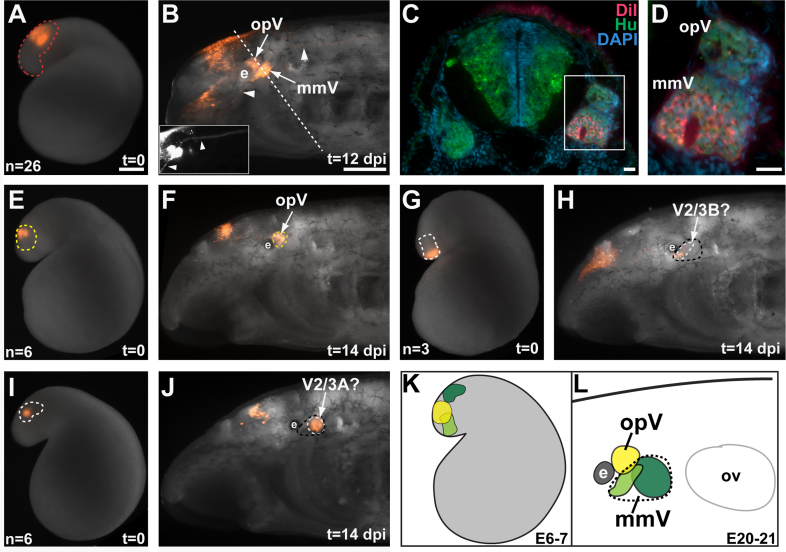
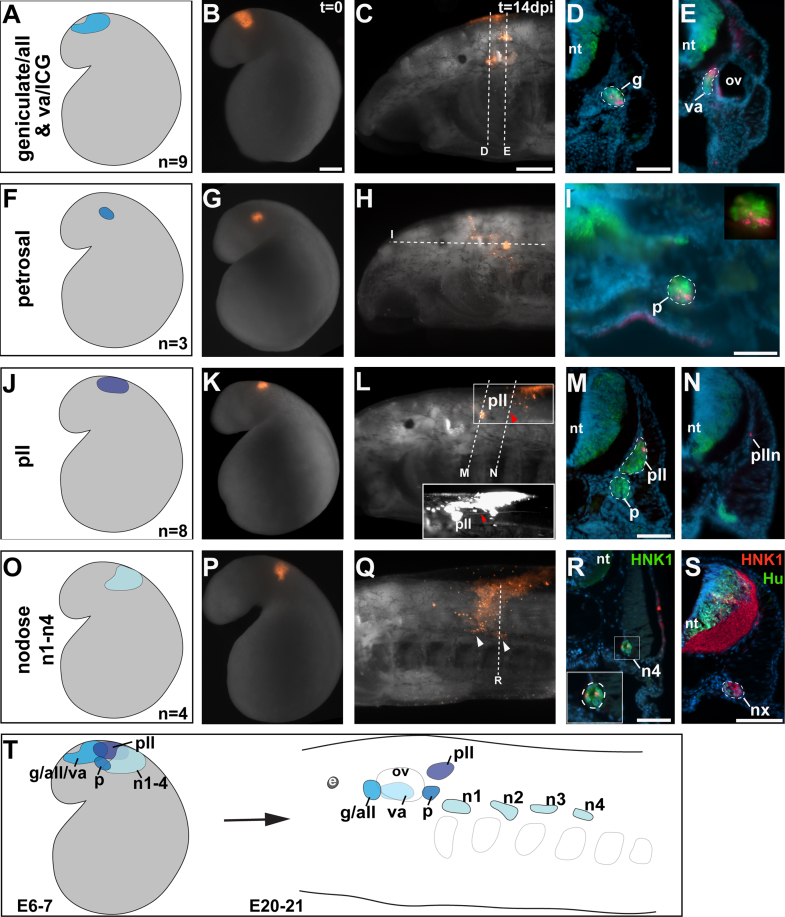
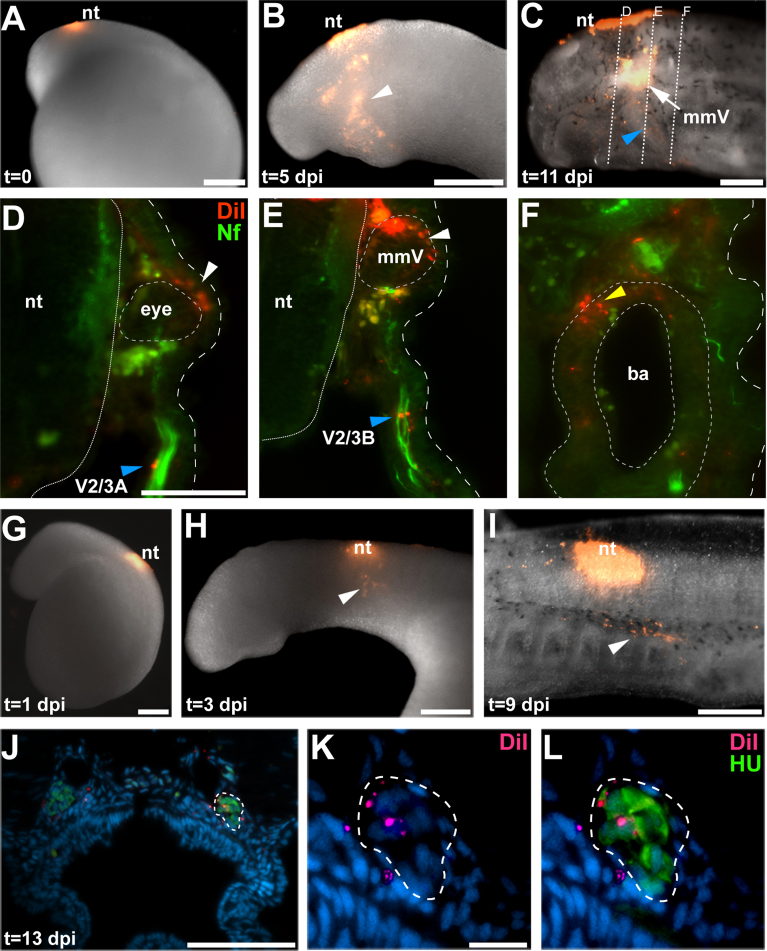
Cranial neurogenic placodes and the neural crest make essential contributions to key adult characteristics of all vertebrates, including the paired peripheral sense organs and craniofacial skeleton. Neurogenic placode development has been extensively characterized in representative jawed vertebrates (gnathostomes) but not in jawless fishes (agnathans). Here, we use in vivo lineage tracing with DiI, together with neuronal differentiation markers, to establish the first detailed fate-map for placode-derived sensory neurons in a jawless fish, the sea lamprey Petromyzon marinus, and to confirm that neural crest cells in the lamprey contribute to the cranial sensory ganglia. We also show that a pan-Pax3/7 antibody labels ophthalmic trigeminal (opV, profundal) placode-derived but not maxillomandibular trigeminal (mmV) placode-derived neurons, mirroring the expression of gnathostome Pax3 and suggesting that Pax3 (and its single Pax3/7 lamprey ortholog) is a pan-vertebrate marker for opV placode-derived neurons. Unexpectedly, however, our data reveal that mmV neuron precursors are located in two separate domains at neurula stages, with opV neuron precursors sandwiched between them. The different branches of the mmV nerve are not comparable between lampreys and gnatho-stomes, and spatial segregation of mmV neuron precursor territories may be a derived feature of lampreys. Nevertheless, maxillary and mandibular neurons are spatially segregated within gnathostome mmV ganglia, suggesting that a more detailed investigation of gnathostome mmV placode development would be worthwhile. Overall, however, our results highlight the conservation of cranial peripheral sensory nervous system development across vertebrates, yielding insight into ancestral vertebrate traits.
Figures
References
-
- Ayer-Le Lièvre C.S., Le Douarin N.M. The early development of cranial sensory ganglia and the potentialities of their component cells studied in quail–chick chimeras. Dev. Biol. 1982;94:291–310. - PubMed
-
- Baker C.V.H., Stark M.R., Marcelle C., Bronner-Fraser M. Competence, specification and induction of Pax-3 in the trigeminal placode. Development. 1999;126:147–156. - PubMed
-
- Barreiro-Iglesias A., Gómez-López M.P., Anadón R., Rodicio M.C. Early development of the cranial nerves in a primitive vertebrate, the sea lamprey, Petromyzon marinus L. Open Zool. J. 2008;1:37–43.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
