

Review
doi: 10.1038/nrg3662.
Epub 2014 Feb 11.
Coupling mRNA processing with transcription in time and space
Affiliations
- PMID: 24514444
- PMCID: PMC4304646
- DOI: 10.1038/nrg3662
Item in Clipboard
Review
Coupling mRNA processing with transcription in time and space
Nat Rev Genet.
2014 Mar.
Abstract
Maturation of mRNA precursors often occurs simultaneously with their synthesis by RNA polymerase II (Pol II). The co-transcriptional nature of mRNA processing has permitted the evolution of coupling mechanisms that coordinate transcription with mRNA capping, splicing, editing and 3' end formation. Recent experiments using sophisticated new methods for analysis of nascent RNA have provided important insights into the relative amount of co-transcriptional and post-transcriptional processing, the relationship between mRNA elongation and processing, and the role of the Pol II carboxy-terminal domain (CTD) in regulating these processes.
Conflict of interest statement
Figures

Human protein names are given throughout. a | The RNA is shown in green; both GTP and the added guanosine cap (Gp) are shown in blue. The mRNA-capping enzyme in metazoans is bifunctional and has both triphosphatase and guanylyl-transferase activities that remove the γ-phosphate of the nascent transcript and transfer GMP from the GTP donor, respectively. The methyl donor S-adenosyl-l -methionine (SAM) is converted to S-adenosyl-l -homocysteine (SAH), which results in the 7-methylguanosine cap (shown in pink). b | Splicing clips out an intron or intervening sequences as a lariat and ligates the flanking exons together through two transesterification reactions. Conserved intronic splicing elements are indicated in red. Spliceosomal U1, U2, U4, U5 and U6 small nuclear ribonucleoprotein particles (snRNPs) and U2 auxiliary factor (U2AF) are shown, but numerous spliceosomal proteins are omitted. c | 3′ ends of mRNAs are formed by coupled cleavage and polyadenylation. Cleavage of mammalian pre-mRNAs occurs ∼25 bases downstream of a consensus sequence (AAUAAA) and is carried out by the multisubunit complex (shown in purple), which comprises cleavage stimulation factor (CstF), cleavage and polyadenylation specificity factor (CPSF) that bears the endonuclease, and cleavage factors I and II (CFIm and CFIIm). Poly(A) polymerase (PAP) adds the poly(A) tail. 3′ ends of non-polyadenylated histone mRNAs (not shown) are also made co-transcriptionally by a cleavage complex that has many subunits in common with CstF and CPSF. The 5′-to-3′ RNA exonuclease 2 (XRN2) degrades RNA downstream of the cleavage site and facilitates transcription termination. d | Adenosine-to-inosine (A-to-I) editing is carried out by adenosine deaminases acting on RNA (ADAR), which deaminate adenosines into inosines. The folded GLUR-2 pre-mRNA substrate is shown with the exon in blue and intron in grey. Pi, inorganic phosphate. Part b is modified with permission from Chen & Cheng (2012) Biosci. Rep.
32, 345-359. © Biochemical Society.
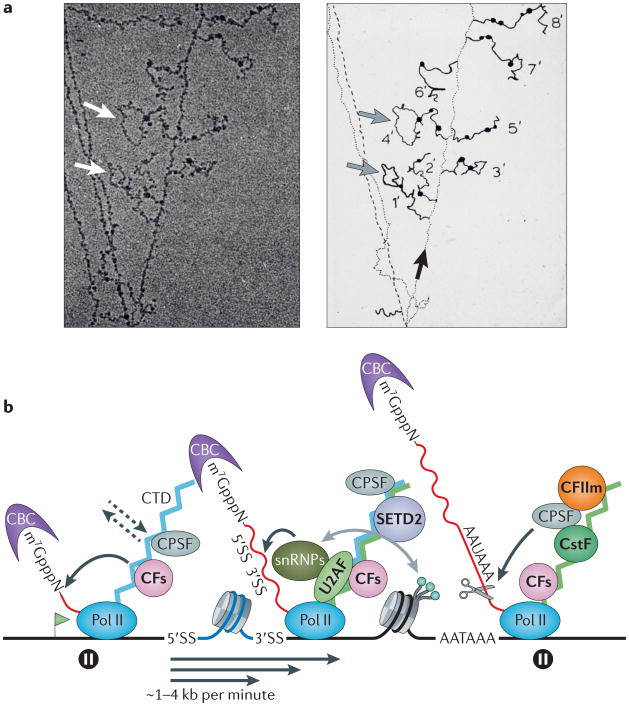
a | Transcription of a gene and co-transcriptional processing of nascent transcripts are shown.‘Miller spread’ electron micrograph (left) and its interpretation (right) are shown for a Drosophila melanogaster embryonic gene. The electron micrograph shows the DNA template with several engaged RNA polymerase II (Pol II) molecules and their associated nascent RNA transcripts with bound proteins (seen as dark blobs) that extend on either side of the DNA. Grey and white arrows mark introns that are spliced out co-transcriptionally. The black arrow indicates the direction of transcription along the DNA template. b | A co-transcriptional mRNA processing scheme is shown. A transcribed gene is shown with multiple polymerases, and the transcription start site is indicated with a green flag. Processing reactions (shown by curved black arrows) are carried out by Pol II-associated factors that act on the nascent transcript (shown in red) at 5′ and 3′ splice sites (5′SS and 3′SS) and the poly(A) site (AAUAAA). Capping factors (CFs, including the mRNA-capping enzyme and guanine-7-methyltransferase) and 3′ end processing/termination factors all bind directly to the carboxy-terminal domain (CTD) of Pol II (but not all factors that bind to the CTD are shown). Transcription elongation (shown by straight black arrows) occurs at variable rates with strong pauses near the transcription start site and downstream of the poly(A) site (shown by pause signs). Proteins that make direct contacts with the CTD are shown in bold, and the dynamic nature of binding is indicated by dashed arrows. CTD phosphorylation changes during the transcription cycle with high levels of phospho-Ser5 (represented by the CTD in blue) at the 5′end (which binds to CFs) and high levels of phospho-Ser2 (represented by the CTD in green) at the 3′end (which binds to the PCF11 subunit of cleavage factor II (CFIIm) (shown in orange)). Note that some 3′end processing factors — including cleavage and polyadenylation specificity factor (CPSF), CFs and the cap-binding complex (CBC) — are present on transcription elongation complexes throughout the length of genes. Nucleosomes that wrap the DNA template have different densities and methylation patterns in exons and introns. Splicing and trimethylation of histone H3 lysine 36 (shown as green balls) by the methyltransferase SET domain-containing 2 (SETD2) may influence one another to establish and maintain splicing patterns (shown by the grey arrow). CstF, cleavage stimulation factor; snRNP, small nuclear ribonucleoparticle; U2AF, U2 auxiliary factor. Part a is reproduced, with permission, from REF. 1 © (1988) Cold Spring Harbor Laboratory Press.
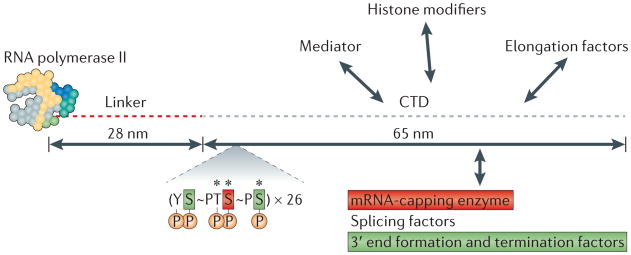
The core yeast RNA polymerase II structure (which consists of ten subunits and has a mass of ∼500 kDa) and its extended carboxy-terminal domain (CTD, which contains 26 heptads with the consensus sequence YSPTSPS) are shown to scale. The human CTD has 52 heptads with the consensus sequence YSPTSPS. The five phosphorylation (P) sites, two peptidyl-prolyl bonds (∼) that are converted between cis and trans conformations and three sites of O-GlcNAcylation (*) are indicated. Heptads with phospho-Ser5 make conserved direct interactions with the mRNA-capping enzyme, whereas heptads with phospho-Ser2 and phospho-Ser7 directly interact with the 3′ end processing/termination factor Pcf11, and the integrator complex, respectively. The figure is modified, with permission, from REF. 188 © (2001) American Association for the Advancement of Science.
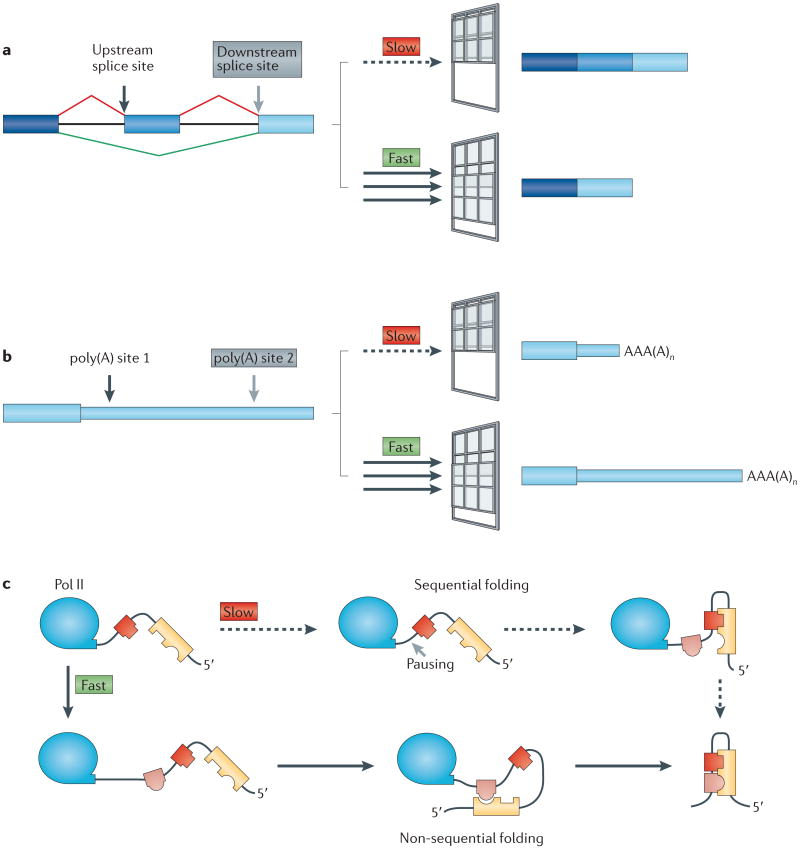
Transcription elongation rate determines the length of the ‘window of opportunity’ for an upstream event to occur on the nascent RNA before it must compete with a downstream event. Slow elongation widens the window of opportunity for commitment to processing at upstream splice sites and poly(A) sites. This could lead to inclusion of alternative exons (part a) and 3′end formation at upstream poly(A) sites (part b), which results in mRNAs with shorter 3′untranslated regions. Slow elongation also favours RNA folding by base-pairing of proximal complementary sequences (represented by interlocking shapes) (part c), which results in sequential rather than non-sequential folding,. Part c is modified, with permission, from REF. 134 © (2006) Annual Reviews.
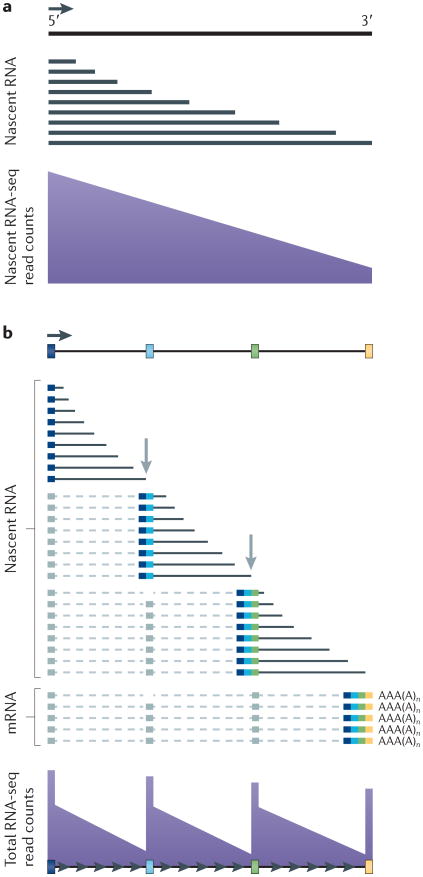
a | Nascent transcripts are mostly incomplete, and 5′ sequences are therefore over-represented relative to 3′ sequences across the gene in nascent RNA sequencing (RNA-seq) profiles. b | Co-transcriptional splicing of nascent transcripts causes a saw-tooth profile in read counts from sequencing of total RNA owing to rapid removal and degradation of sequences from the 3′ ends of introns shortly after the 3′ splice site has been transcribed. Figure is modified, with permission, from REF. 74 © (2011) Macmillian Publishers Ltd. All rights reserved.
References
-
- Beyer AL, Osheim YN. Splice site selection, rate of splicing, and alternative splicing on nascent transcripts. Genes Dev. 1988;2:754–765. This is an early graphic demonstration of co-transcriptional splicing. - PubMed
-
- Bieberstein NI, Carrillo Oesterreich F, Straube K, Neugebauer KM. First exon length controls active chromatin signatures and transcription. Cell Rep. 2012;2:62–68. - PubMed
-
- Gomez Acuna LI, Fiszbein A, Allo M, Schor IE, Kornblihtt AR. Connections between chromatin signatures and splicing. Wiley Interdiscip Rev RNA. 2013;4:77–91. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
