

3'-nucleotidase/nuclease activity allows Leishmania parasites to escape killing by neutrophil extracellular traps
- PMID: 24516114
- PMCID: PMC3993383
- DOI: 10.1128/IAI.01232-13
3'-nucleotidase/nuclease activity allows Leishmania parasites to escape killing by neutrophil extracellular traps
Abstract
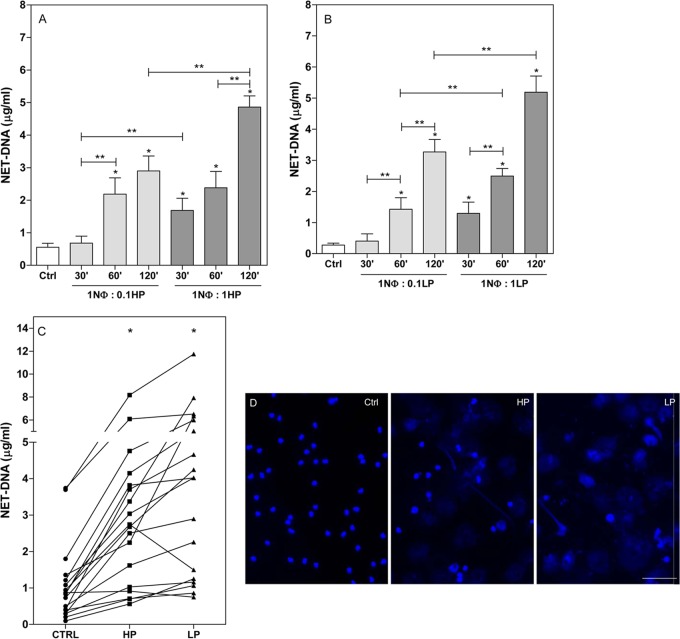
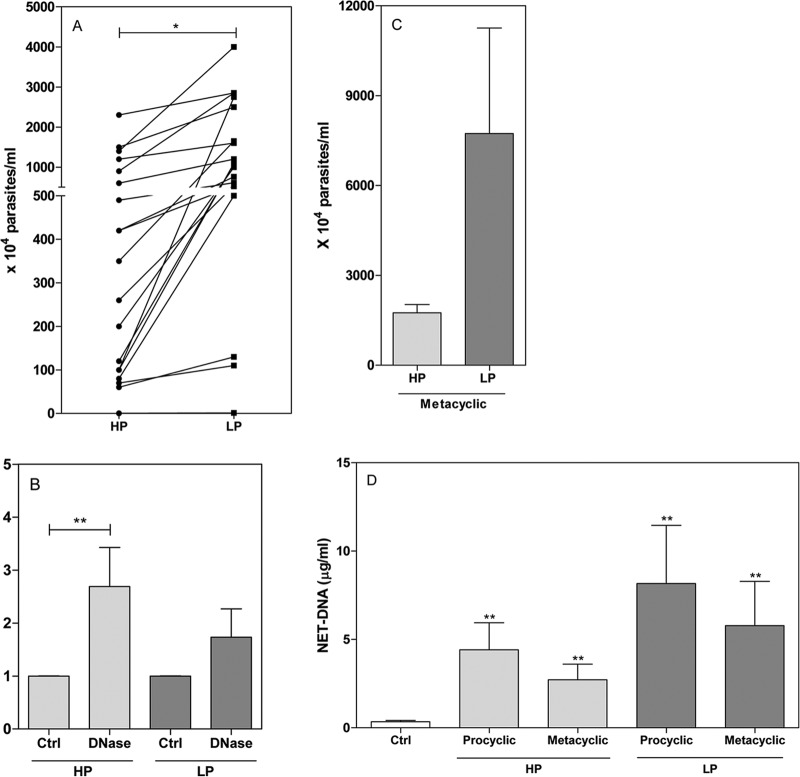
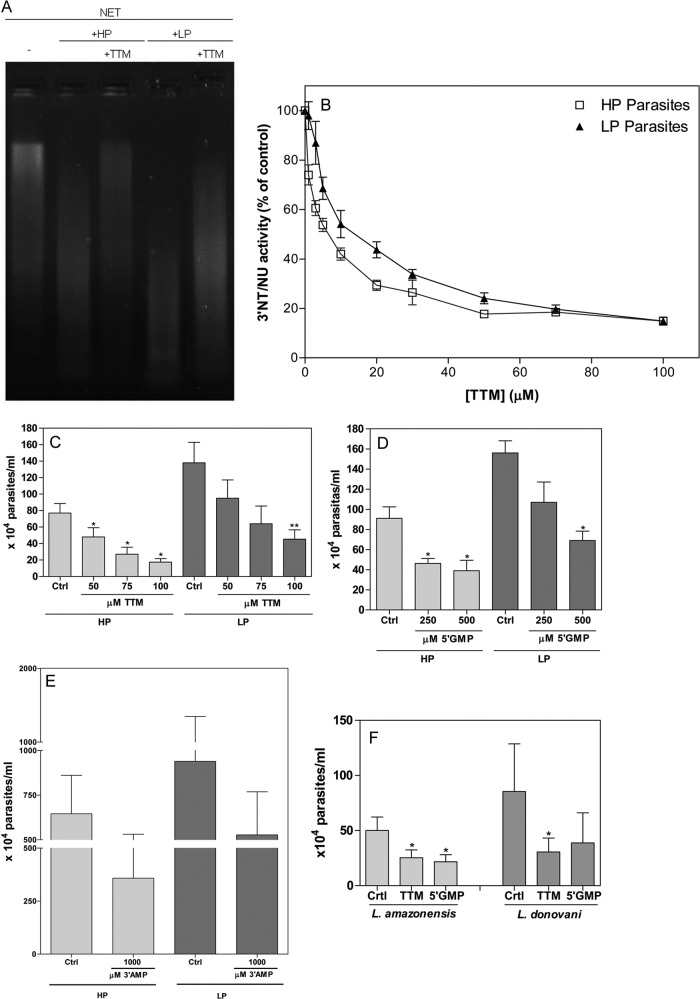
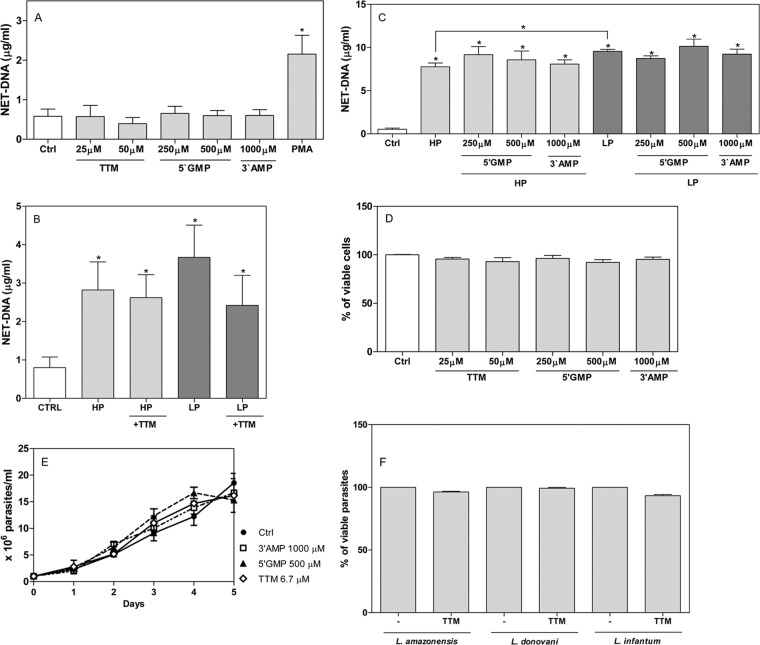
Leishmaniasis is a widespread neglected tropical disease caused by parasites of the Leishmania genus. These parasites express the enzyme 3'-nucleotidase/nuclease (3'NT/NU), which has been described to be involved in parasite nutrition and infection. Bacteria that express nucleases escape the toxic effects of neutrophil extracellular traps (NETs). Hence, we investigated the role of 3'NT/NU in Leishmania survival of NET-mediated killing. Promastigotes of Leishmania infantum were cultured in high-phosphate (HP) or low-phosphate (LP) medium to modulate nuclease activity. We compared the survival of the two different groups of Leishmania during interaction with human neutrophils, assessing the role of neutrophil extracellular traps. As previously reported, we detected higher nuclease activity in parasites cultured in LP medium. Both LP and HP promastigotes were capable of inducing the release of neutrophil extracellular traps from human neutrophils in a dose- and time-dependent manner. LP parasites had 2.4 times more survival than HP promastigotes. NET disruption was prevented by the treatment of the parasites with ammonium tetrathiomolybdate (TTM), a 3'NT/NU inhibitor. Inhibition of 3'NT/NU by 3'-AMP, 5'-GMP, or TTM decreased promastigote survival upon interaction with neutrophils. Our results show that Leishmania infantum induces NET release and that promastigotes can escape NET-mediated killing by 3'-nucleotidase/nuclease activity, thus ascribing a new function to this enzyme.
Figures
Similar articles
-
Cloning, expression and purification of 3'-nucleotidase/nuclease, an enzyme responsible for the Leishmania escape from neutrophil extracellular traps.Mol Biochem Parasitol. 2019 Apr;229:6-14. doi: 10.1016/j.molbiopara.2019.02.004. Epub 2019 Feb 14. Mol Biochem Parasitol. 2019. PMID: 30772424
-
Candida Extracellular Nucleotide Metabolism Promotes Neutrophils Extracellular Traps Escape.Front Cell Infect Microbiol. 2021 Jul 13;11:678568. doi: 10.3389/fcimb.2021.678568. eCollection 2021. Front Cell Infect Microbiol. 2021. PMID: 34327150 Free PMC article.
-
3'nucleotidase/nuclease in protozoan parasites: Molecular and biochemical properties and physiological roles.Exp Parasitol. 2017 Aug;179:1-6. doi: 10.1016/j.exppara.2017.06.001. Epub 2017 Jun 3. Exp Parasitol. 2017. PMID: 28587841 Review.
-
Inhibitory effects promoted by 5'-nucleotides on the ecto-3'-nucleotidase activity of Leishmania amazonensis.Exp Parasitol. 2016 Oct;169:111-8. doi: 10.1016/j.exppara.2016.08.001. Epub 2016 Aug 13. Exp Parasitol. 2016. PMID: 27531705
-
Understanding the genetic complexity of Leishmania infantum in the Americas: a focus on 3'NT/NU gene deletion.Mem Inst Oswaldo Cruz. 2025 Mar 24;120:e240160. doi: 10.1590/0074-02760240160. eCollection 2025. Mem Inst Oswaldo Cruz. 2025. PMID: 40136219 Free PMC article. Review.
Cited by
-
Far beyond Phagocytosis: Phagocyte-Derived Extracellular Traps Act Efficiently against Protozoan Parasites In Vitro and In Vivo.Mediators Inflamm. 2016;2016:5898074. doi: 10.1155/2016/5898074. Epub 2016 Jun 30. Mediators Inflamm. 2016. PMID: 27445437 Free PMC article. Review.
-
Extracellular Traps Released by Neutrophils from Cats are Detrimental to Toxoplasma gondii Infectivity.Microorganisms. 2020 Oct 22;8(11):1628. doi: 10.3390/microorganisms8111628. Microorganisms. 2020. PMID: 33105542 Free PMC article.
-
Dependence of Leishmania parasite on host derived ATP: an overview of extracellular nucleotide metabolism in parasite.J Parasit Dis. 2019 Mar;43(1):1-13. doi: 10.1007/s12639-018-1061-4. Epub 2018 Dec 1. J Parasit Dis. 2019. PMID: 30956439 Free PMC article. Review.
-
Revisiting the Mechanisms of Immune Evasion Employed by Human Parasites.Front Cell Infect Microbiol. 2021 Jul 29;11:702125. doi: 10.3389/fcimb.2021.702125. eCollection 2021. Front Cell Infect Microbiol. 2021. PMID: 34395313 Free PMC article. Review.
-
Evasion of Neutrophil Extracellular Traps by Respiratory Pathogens.Am J Respir Cell Mol Biol. 2017 Apr;56(4):423-431. doi: 10.1165/rcmb.2016-0193PS. Am J Respir Cell Mol Biol. 2017. PMID: 27854516 Free PMC article. Review.
References
-
- Urban CF, Ermert D, Schmid M, Abu-Abed U, Goosmann C, Nacken W, Brinkmann V, Jungblut PR, Zychlinsky A. 2009. Neutrophil extracellular traps contain calprotectin, a cytosolic protein complex involved in host defense against Candida albicans. PLoS Pathog. 5:e1000639. 10.1371/journal.ppat.1000639 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
