

Promiscuous MYC locus rearrangements hijack enhancers but mostly super-enhancers to dysregulate MYC expression in multiple myeloma
- PMID: 24518206
- PMCID: PMC4126852
- DOI: 10.1038/leu.2014.70
Promiscuous MYC locus rearrangements hijack enhancers but mostly super-enhancers to dysregulate MYC expression in multiple myeloma
Abstract
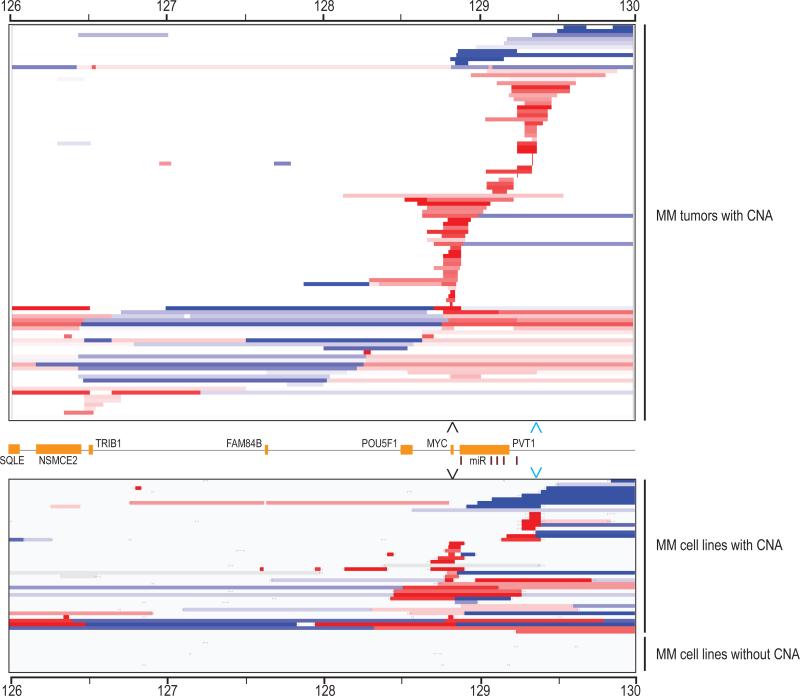
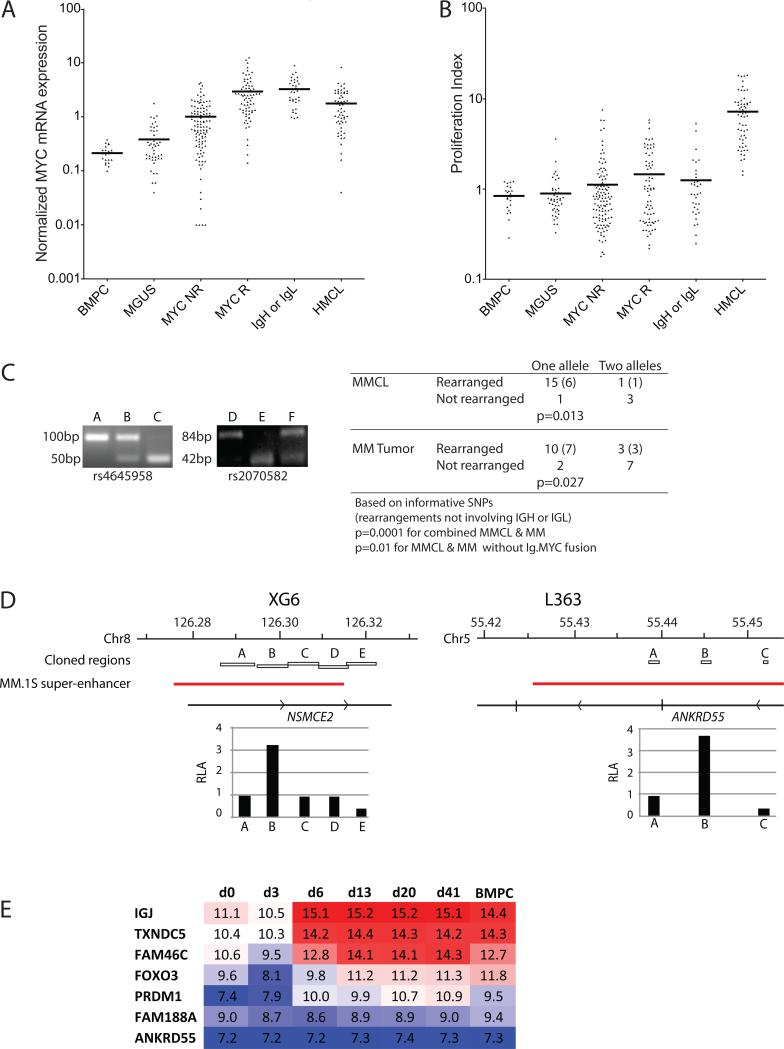
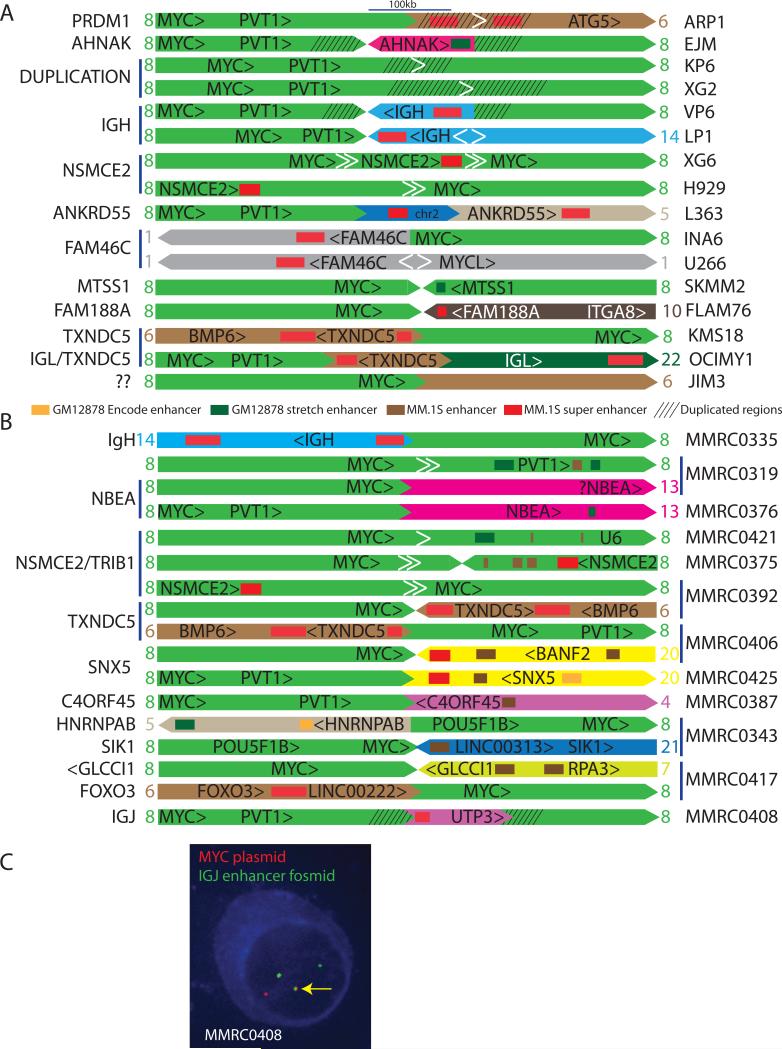
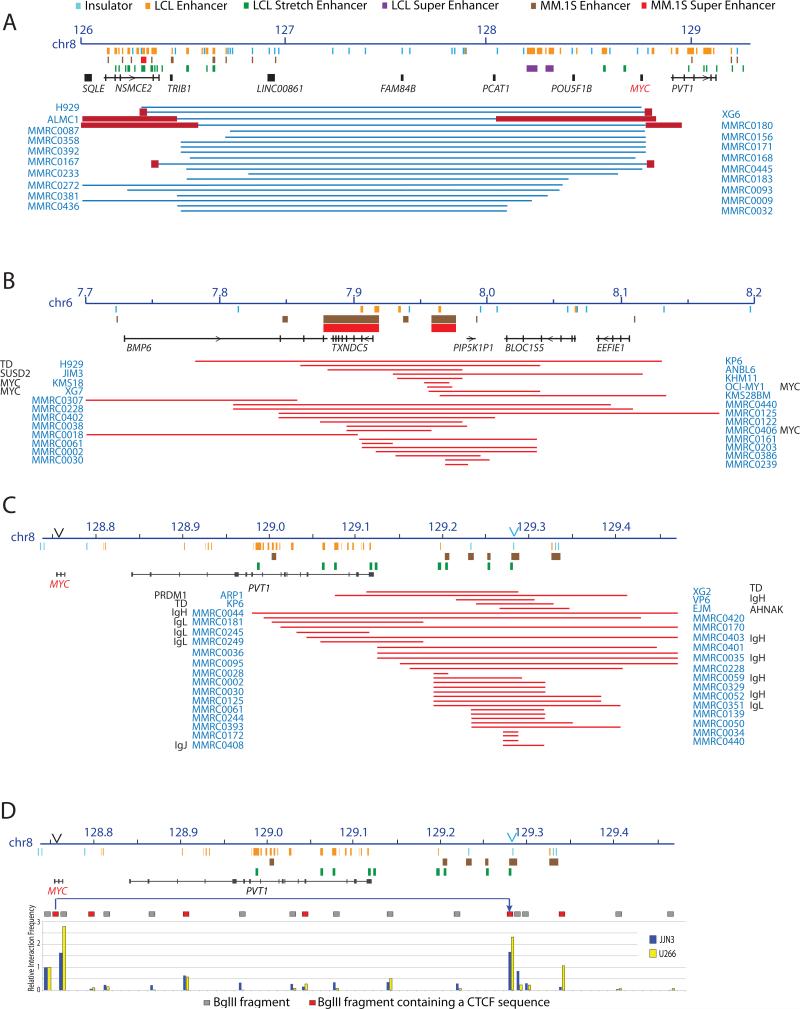
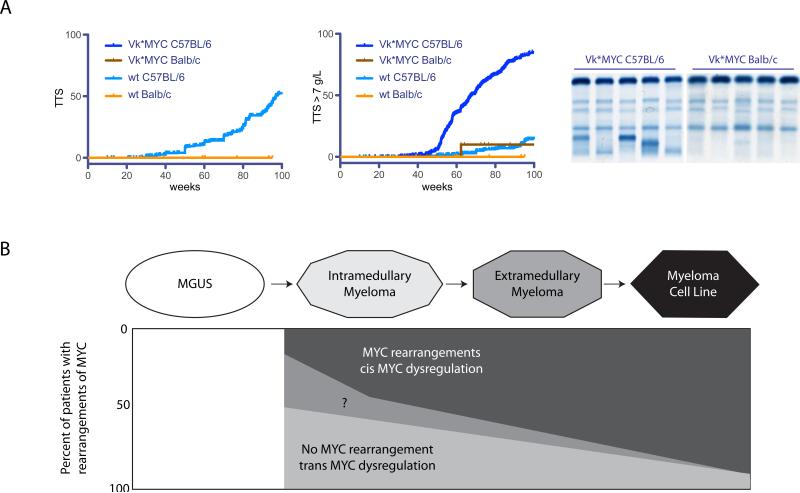
MYC locus rearrangements-often complex combinations of translocations, insertions, deletions and inversions-in multiple myeloma (MM) were thought to be a late progression event, which often did not involve immunoglobulin genes. Yet, germinal center activation of MYC expression has been reported to cause progression to MM in an MGUS (monoclonal gammopathy of undetermined significance)-prone mouse strain. Although previously detected in 16% of MM, we find MYC rearrangements in nearly 50% of MM, including smoldering MM, and they are heterogeneous in some cases. Rearrangements reposition MYC near a limited number of genes associated with conventional enhancers, but mostly with super-enhancers (e.g., IGH, IGL, IGK, NSMCE2, TXNDC5, FAM46C, FOXO3, IGJ, PRDM1). MYC rearrangements are associated with a significant increase of MYC expression that is monoallelic, but MM tumors lacking a rearrangement have biallelic MYC expression at significantly higher levels than in MGUS. We also have shown that germinal center activation of MYC does not cause MM in a mouse strain that rarely develops spontaneous MGUS. It appears that increased MYC expression at the MGUS/MM transition usually is biallelic, but sometimes can be monoallelic if there is an MYC rearrangement. Our data suggest that MYC rearrangements, regardless of when they occur during MM pathogenesis, provide one event that contributes to tumor autonomy.
Figures
References
-
- Dalla-Favera R, Martinotti S, Gallo RC, Erikson J, Croce CM. Translocation and rearrangements of the c-myc oncogene locus in human undifferentiated B-cell lymphomas. Science. 1983 Feb 25;219(4587):963–967. - PubMed
-
- Potter M. Neoplastic development in plasma cells. Immunol Rev. 2003 Aug;194:177–195. - PubMed
-
- Kuppers R. Mechanisms of B-cell lymphoma pathogenesis. Nat Rev Cancer. 2005 Apr;5(4):251–262. - PubMed
-
- Zhan F, Tian E, Bumm K, Smith R, Barlogie B, Shaughnessy J., Jr. Gene expression profiling of human plasma cell differentiation and classification of multiple myeloma 23 based on similarities to distinct stages of late-stage B-cell development. Blood. 2003 Feb 1;101(3):1128–1140. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
