

Host-specific adaptation of HIV-1 subtype B in the Japanese population
- PMID: 24522911
- PMCID: PMC3993807
- DOI: 10.1128/JVI.00147-14
Host-specific adaptation of HIV-1 subtype B in the Japanese population
Abstract
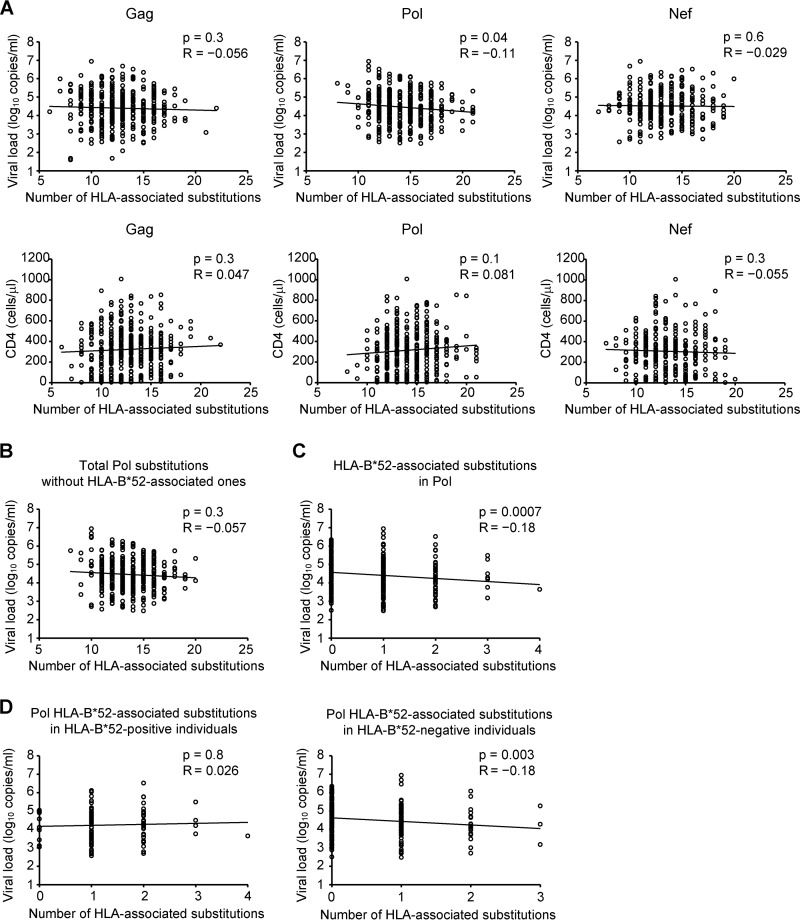
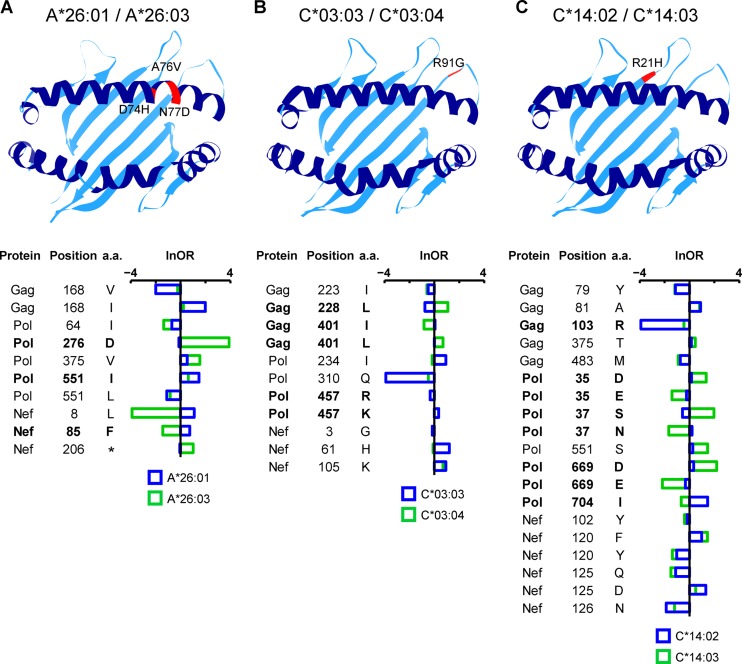
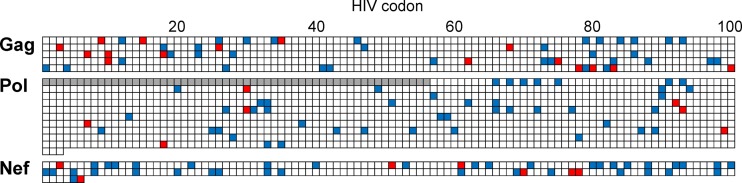
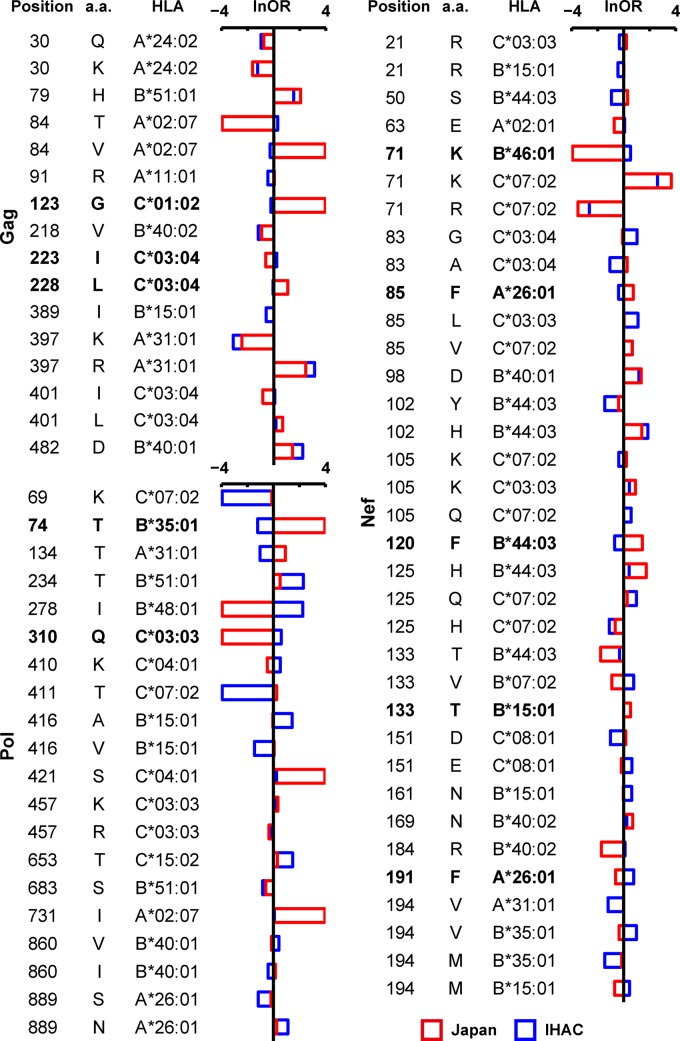
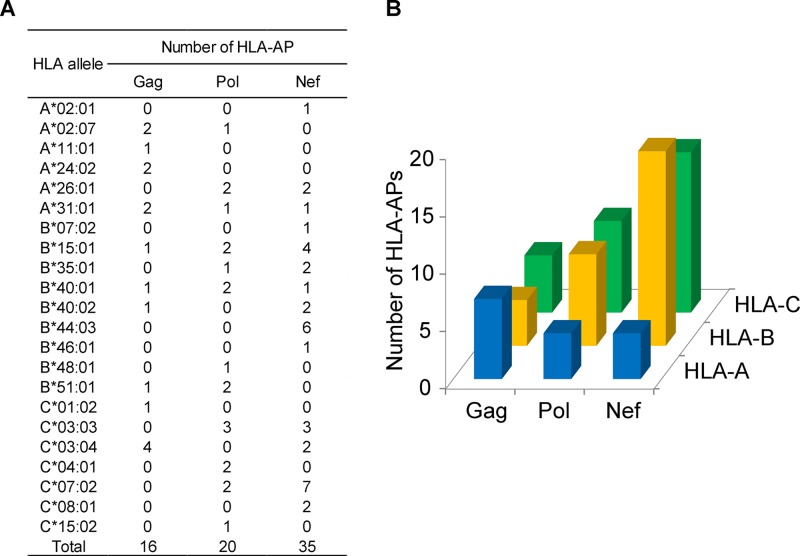
The extent to which HIV-1 clade B strains exhibit population-specific adaptations to host HLA alleles remains incompletely known, in part due to incomplete characterization of HLA-associated HIV-1 polymorphisms (HLA-APs) in different global populations. Moreover, it remains unknown to what extent the same HLA alleles may drive significantly different escape pathways across populations. As the Japanese population exhibits distinctive HLA class I allele distributions, comparative analysis of HLA-APs between HIV-1 clade B-infected Japanese and non-Asian cohorts could shed light on these questions. However, HLA-APs remain incompletely mapped in Japan. In a cohort of 430 treatment-naive Japanese with chronic HIV-1 clade B infection, we identified 284 HLA-APs in Gag, Pol, and Nef using phylogenetically corrected methods. The number of HLA-associated substitutions in Pol, notably those restricted by HLA-B*52:01, was weakly inversely correlated with the plasma viral load (pVL), suggesting that the transmission and persistence of B*52:01-driven Pol mutations could modulate the pVL. Differential selection of HLA-APs between HLA subtype members, including those differing only with respect to substitutions outside the peptide-binding groove, was observed, meriting further investigation as to their mechanisms of selection. Notably, two-thirds of HLA-APs identified in Japan had not been reported in previous studies of predominantly Caucasian cohorts and were attributable to HLA alleles unique to, or enriched in, Japan. We also identified 71 cases where the same HLA allele drove significantly different escape pathways in Japan versus predominantly Caucasian cohorts. Our results underscore the distinct global evolution of HIV-1 clade B as a result of host population-specific cellular immune pressures.
Importance: Cytotoxic T lymphocyte (CTL) escape mutations in HIV-1 are broadly predictable based on the HLA class I alleles expressed by the host. Because HLA allele distributions differ among worldwide populations, the pattern and diversity of HLA-associated escape mutations are likely to be somewhat distinct to each race and region. HLA-associated polymorphisms (HLA-APs) in HIV-1 have previously been identified at the population level in European, North American, Australian, and African cohorts; however, large-scale analyses of HIV-1 clade B-specific HLA-APs in Asians are lacking. Differential intraclade HIV-1 adaptation to global populations can be investigated via comparative analyses of HLA-associated polymorphisms across ethnic groups, but such studies are rare. Here, we identify HLA-APs in a large Japanese HIV-1 clade B cohort using phylogenetically informed methods and observe that the majority of them had not been previously characterized in predominantly Caucasian populations. The results highlight HIV's unique adaptation to cellular immune pressures imposed by different global populations.
Figures

References
-
- Brumme ZL, Brumme CJ, Heckerman D, Korber BT, Daniels M, Carlson J, Kadie C, Bhattacharya T, Chui C, Szinger J, Mo T, Hogg RS, Montaner JS, Frahm N, Brander C, Walker BD, Harrigan PR. 2007. Evidence of differential HLA class I-mediated viral evolution in functional and accessory/regulatory genes of HIV-1. PLoS Pathog. 3:e94. 10.1371/journal.ppat.0030094 - DOI - PMC - PubMed
-
- Brumme ZL, John M, Carlson JM, Brumme CJ, Chan D, Brockman MA, Swenson LC, Tao I, Szeto S, Rosato P, Sela J, Kadie CM, Frahm N, Brander C, Haas DW, Riddler SA, Haubrich R, Walker BD, Harrigan PR, Heckerman D, Mallal S. 2009. HLA-associated immune escape pathways in HIV-1 subtype B Gag, Pol and Nef proteins. PLoS One 4:e6687. 10.1371/journal.pone.0006687 - DOI - PMC - PubMed
-
- Brumme ZL, Tao I, Szeto S, Brumme CJ, Carlson JM, Chan D, Kadie C, Frahm N, Brander C, Walker B, Heckerman D, Harrigan PR. 2008. Human leukocyte antigen-specific polymorphisms in HIV-1 Gag and their association with viral load in chronic untreated infection. AIDS 22:1277–1286. 10.1097/QAD.0b013e3283021a8c - DOI - PubMed
-
- Rolland M, Carlson JM, Manocheewa S, Swain JV, Lanxon-Cookson E, Deng W, Rousseau CM, Raugi DN, Learn GH, Maust BS, Coovadia H, Ndung'u T, Goulder PJ, Walker BD, Brander C, Heckerman DE, Mullins JI. 2010. Amino-acid co-variation in HIV-1 Gag subtype C: HLA-mediated selection pressure and compensatory dynamics. PLoS One 5:e12463. 10.1371/journal.pone.0012463 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
