

Trans-signaling is a dominant mechanism for the pathogenic actions of interleukin-6 in the brain
- PMID: 24523541
- PMCID: PMC6802757
- DOI: 10.1523/JNEUROSCI.2830-13.2014
Trans-signaling is a dominant mechanism for the pathogenic actions of interleukin-6 in the brain
Abstract
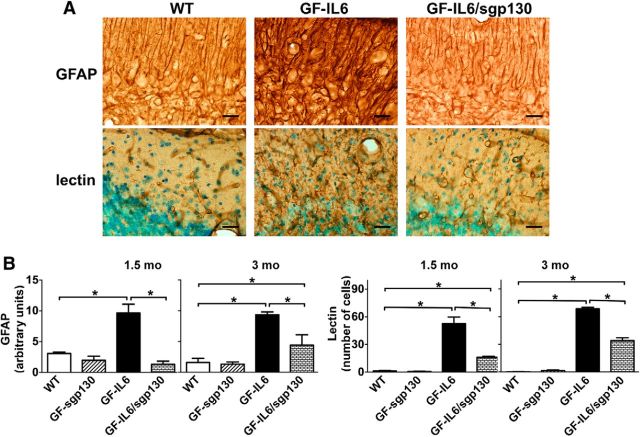
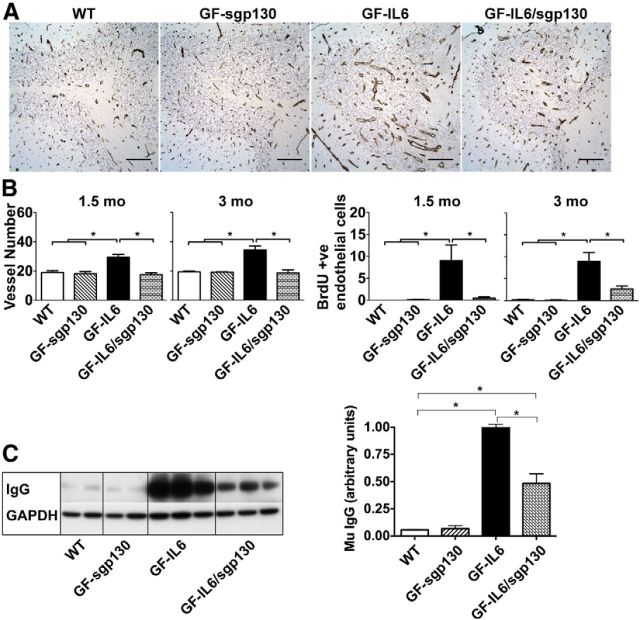
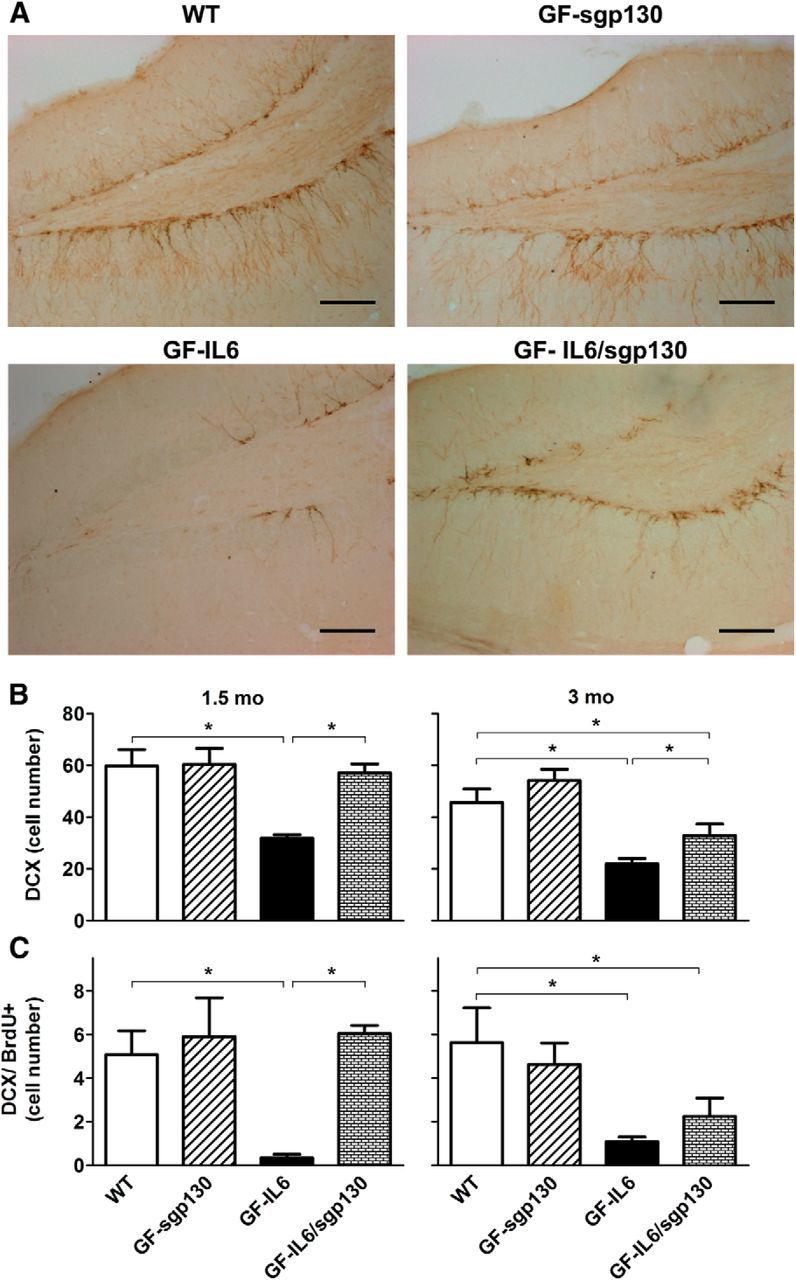
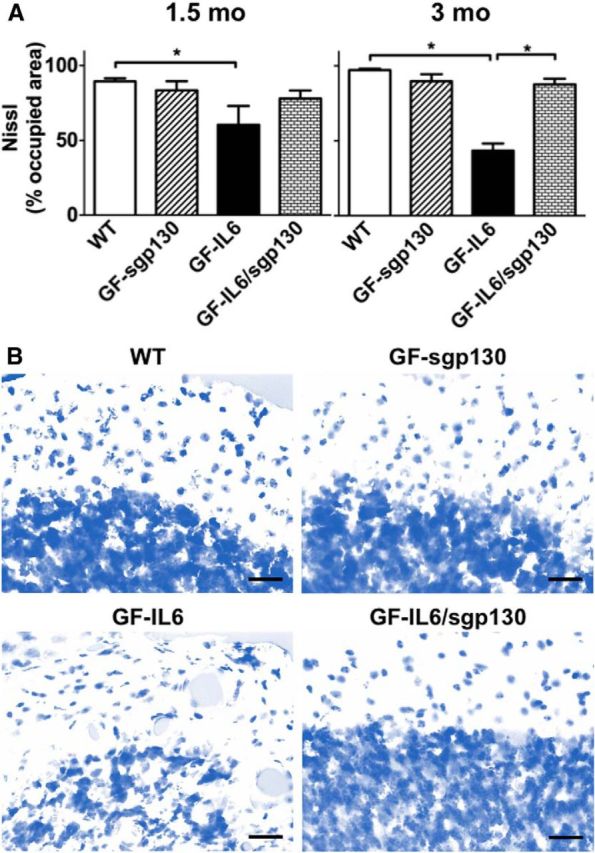
IL-6 is implicated in the pathogenesis of various neuroinflammatory and neurodegenerative disorders of the CNS. IL-6 signals via binding to either the membrane bound IL-6Rα (classic signaling) or soluble (s)IL-6Ra (trans-signaling) that then form a complex with gp130 to activate the JAK/STAT signaling pathway. The importance of classic versus trans-signaling in mediating IL-6 actions in the living CNS is relatively unknown and was the focus of this investigation. Bigenic mice (termed GFAP-IL6/sgp130 mice) were generated with CNS-restricted, astrocyte-targeted production of IL-6 and coproduction of the specific inhibitor of IL-6 trans-signaling, human sgp130-Fc. Transgene-encoded IL-6 mRNA levels were similar in the brain of GFAP-IL6 and GFAP-IL6/sgp130 mice. However, GFAP-IL6/sgp130 mice had decreased pY(705)-STAT3 in the brain due to a reduction in the total number of pY(705)-STAT3-positive cells and a marked loss of pY(705)-STAT3 in specific cell types. Blockade of trans-signaling in the brain of the GFAP-IL6 mice significantly attenuated Serpina3n but not SOCS3 gene expression, whereas vascular changes including angiogenesis and blood-brain barrier leakage as well as gliosis were also reduced significantly. Hippocampal neurogenesis which was impaired in GFAP-IL6 mice was rescued in young GFAP-IL6 mice with cerebral sgp130 production. Finally, degenerative changes in the cerebellum characteristic of GFAP-IL6 mice were absent in GFAP-IL6/sgp130 mice. The findings indicate that in the CNS: (1) sgp130 is able to block IL-6 trans-signaling, (2) trans-signaling is important for IL-6 cellular communication with selective cellular and molecular targets, and (3) blocking of trans-signaling alleviates many of the detrimental effects of IL-6.
Keywords: interleukin-6; neuropathology; signal transduction; trans-signaling; transgenic mouse.
Figures
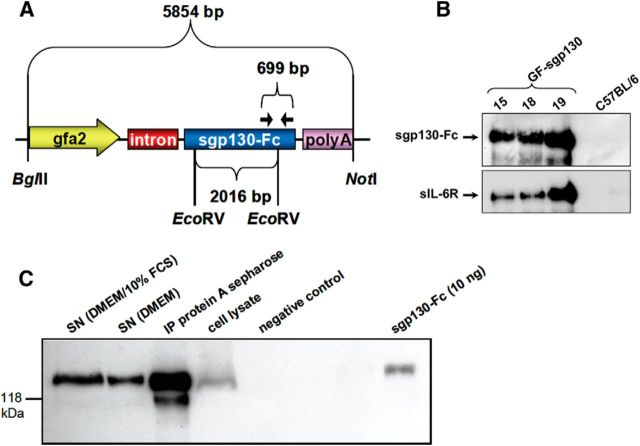
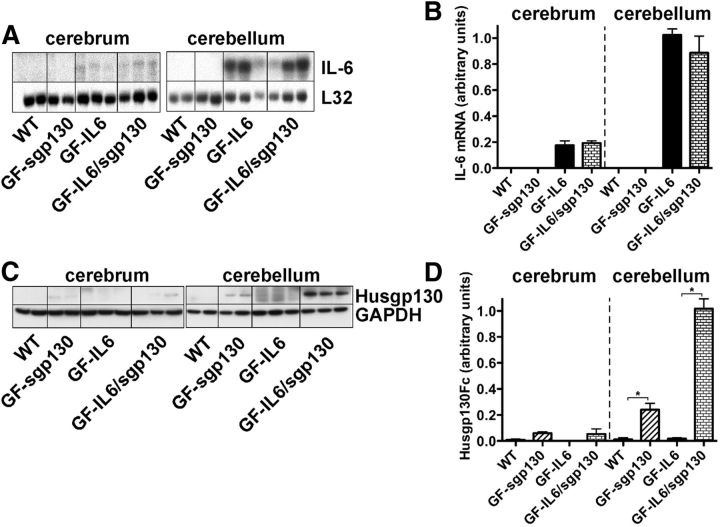
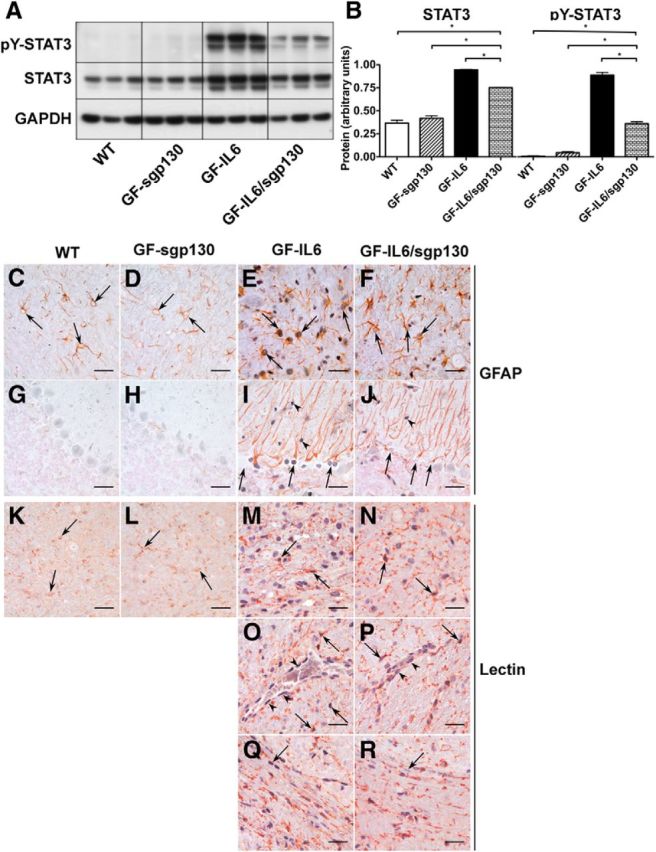
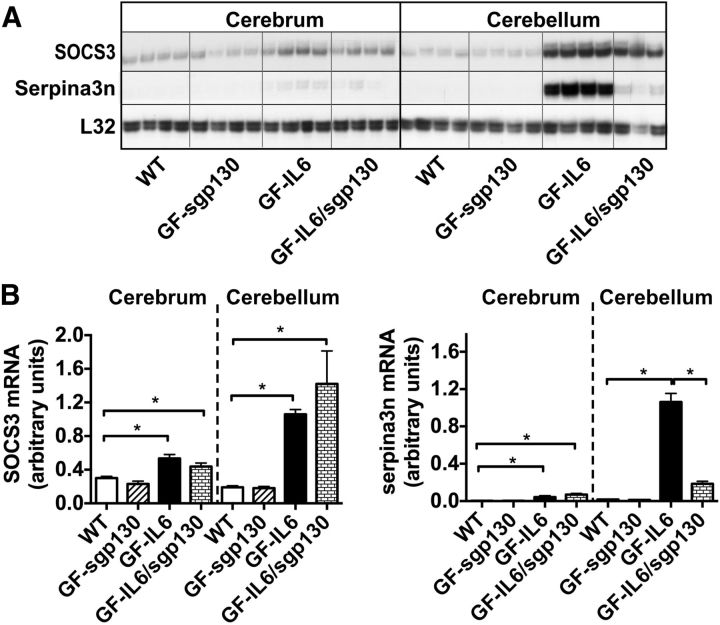
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous