

Small GTPase RhoE/Rnd3 is a critical regulator of Notch1 signaling
- PMID: 24525741
- PMCID: PMC4031027
- DOI: 10.1158/0008-5472.CAN-12-0452
Small GTPase RhoE/Rnd3 is a critical regulator of Notch1 signaling
Abstract
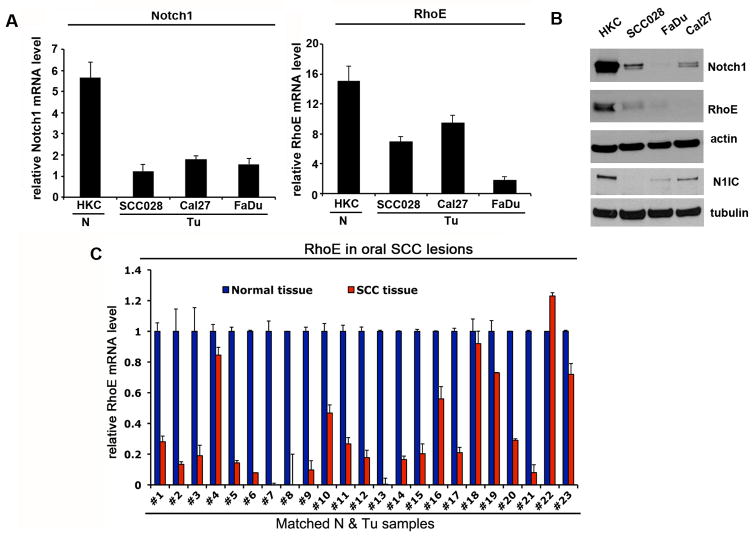
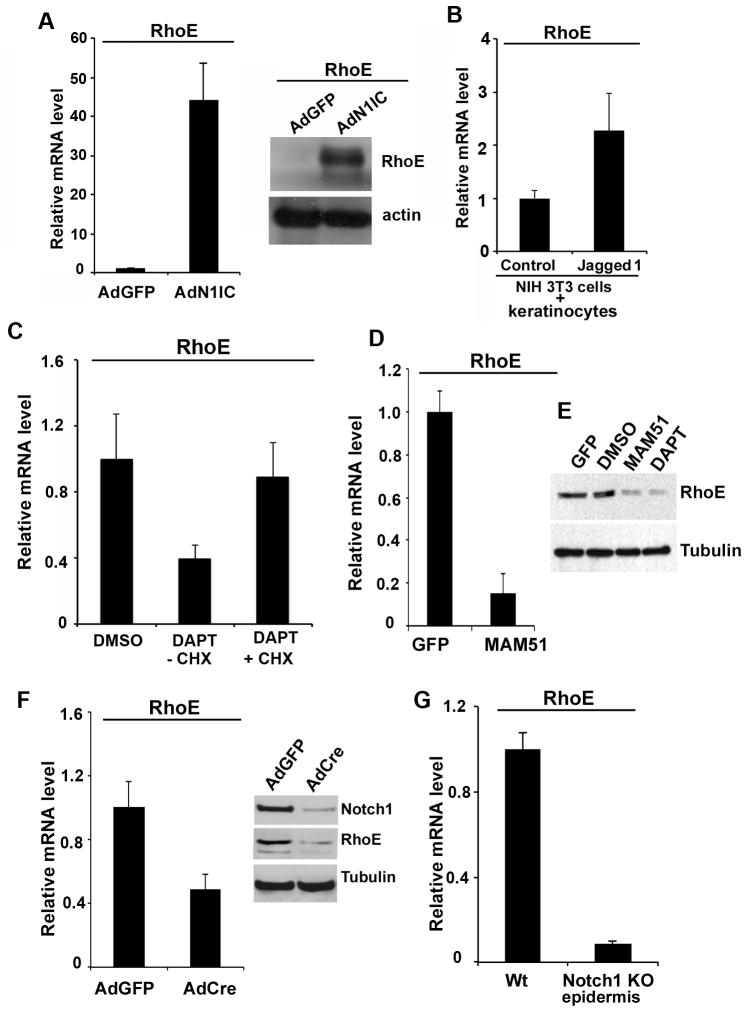
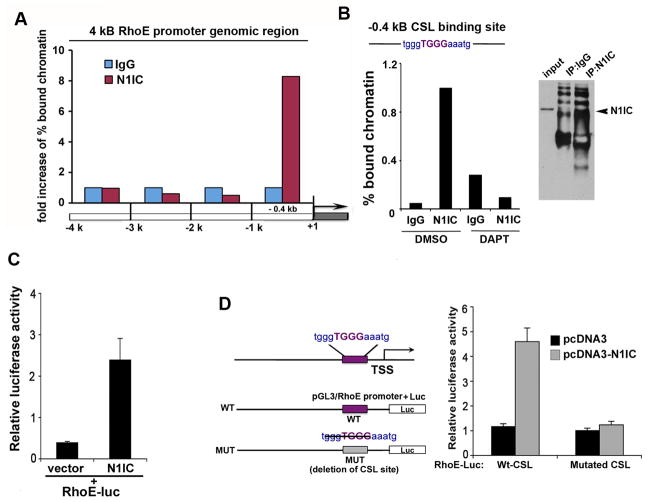
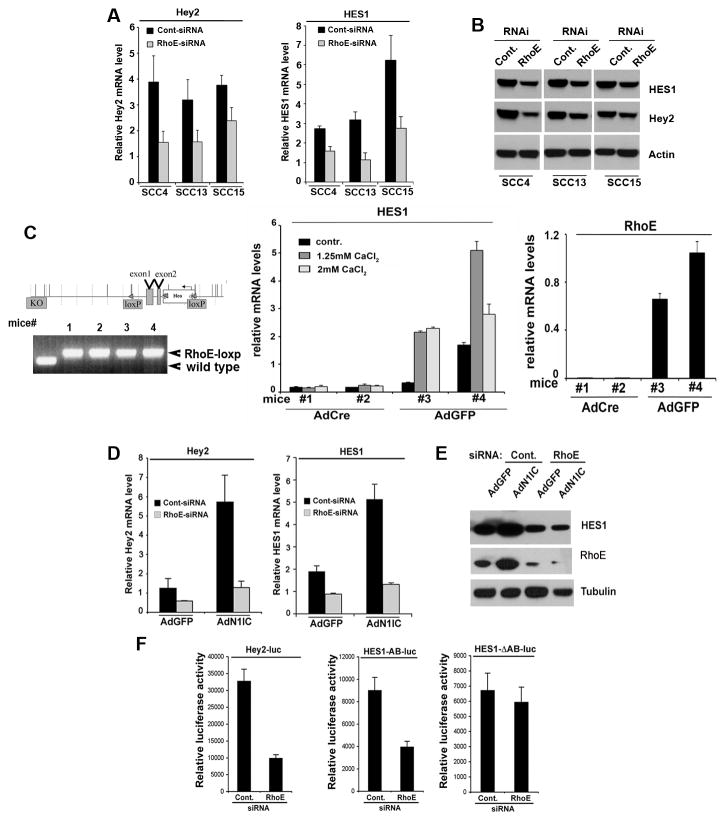
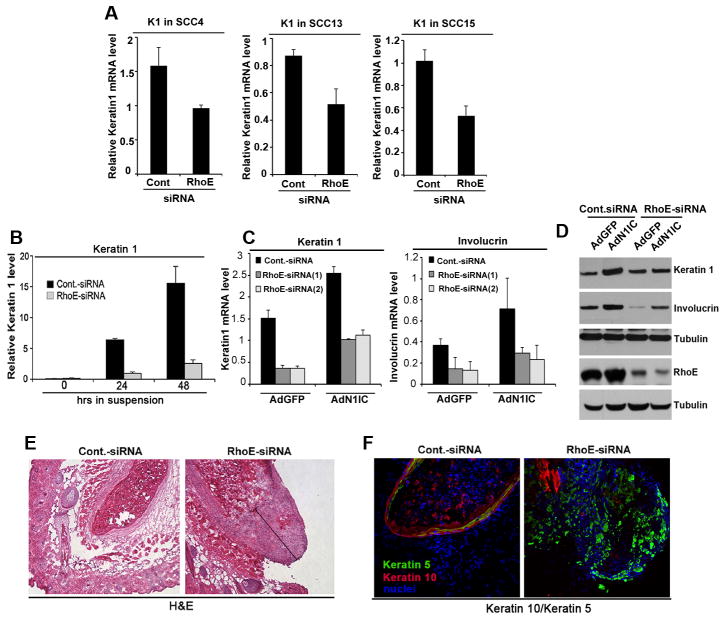
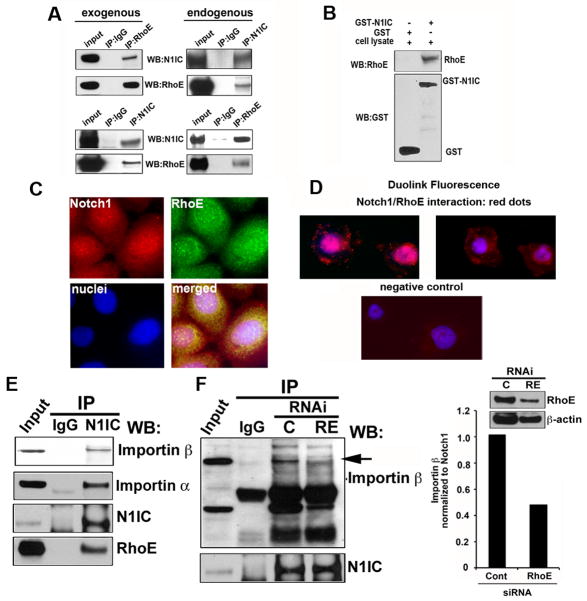
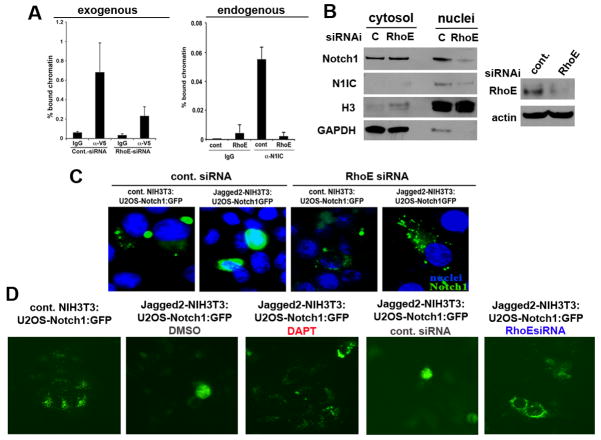
Aberrations of Notch signaling have been implicated in a variety of human cancers. Oncogenic mutations in NOTCH1 are common in human T-cell leukemia and lymphomas. However, loss-of-function somatic mutations in NOTCH1 arising in solid tumors imply a tumor suppressor function, which highlights the need to understand Notch signaling more completely. Here, we describe the small GTPase RhoE/Rnd3 as a downstream mediator of Notch signaling in squamous cell carcinomas (SCC) that arise in skin epithelia. RhoE is a transcriptional target of activated Notch1, which is attenuated broadly in SCC cells. RhoE depletion suppresses Notch1-mediated signaling in vitro, rendering primary keratinocytes resistant to Notch1-mediated differentiation and thereby favoring a proliferative cell fate. Mechanistic investigations indicated that RhoE controls a key step in Notch1 signaling by mediating nuclear translocation of the activated portion of Notch1 (N1IC) through interaction with importins. Our results define RhoE as a Notch1 target that is essential for recruitment of N1IC to the promoters of Notch1 target genes, establishing a regulatory feedback loop in Notch1 signaling. This molecular circuitry may inform distinct cell fate decisions to Notch1 in epithelial tissues, where carcinomas such as SCC arise.
©2014 AACR
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- AR39190/AR/NIAMS NIH HHS/United States
- CA97216/CA/NCI NIH HHS/United States
- CA16038/CA/NCI NIH HHS/United States
- R01 CA073796/CA/NCI NIH HHS/United States
- R01 AR064786/AR/NIAMS NIH HHS/United States
- R01 CA097216/CA/NCI NIH HHS/United States
- R01 AR039190/AR/NIAMS NIH HHS/United States
- CA73796/CA/NCI NIH HHS/United States
- R01 CA140615/CA/NCI NIH HHS/United States
- CA140615/CA/NCI NIH HHS/United States
- CA127247,/CA/NCI NIH HHS/United States
- P01 CA080058/CA/NCI NIH HHS/United States
- P01 CA016038/CA/NCI NIH HHS/United States
- ImNIH/Intramural NIH HHS/United States
- R01 CA127247/CA/NCI NIH HHS/United States
- CA80058/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
