

T-cell responses to oncogenic merkel cell polyomavirus proteins distinguish patients with merkel cell carcinoma from healthy donors
- PMID: 24526738
- PMCID: PMC3994405
- DOI: 10.1158/1078-0432.CCR-13-2697
T-cell responses to oncogenic merkel cell polyomavirus proteins distinguish patients with merkel cell carcinoma from healthy donors
Abstract
Purpose: Merkel cell carcinoma (MCC) is a highly aggressive skin cancer with strong evidence of viral carcinogenesis. The association of MCC with the Merkel cell polyomavirus (MCPyV) may explain the explicit immunogenicity of MCC. Indeed, MCPyV-encoded proteins are likely targets for cytotoxic immune responses to MCC as they are both foreign to the host and necessary to maintain the oncogenic phenotype. However, to date only a single MCPyV-derived CD8 T-cell epitope has been described, thus impeding specific monitoring of T-cell responses to MCC.
Method: To overcome this limitation, we scanned the MCPyV oncoprotein large T and small T antigens and the virus capsid protein VP1 for potential T-cell epitopes, and tested for MHC class I affinity. We confirmed the relevance of these epitopes using a high-throughput platform for T-cell enrichment and combinatorial encoding of MHC class I multimers.
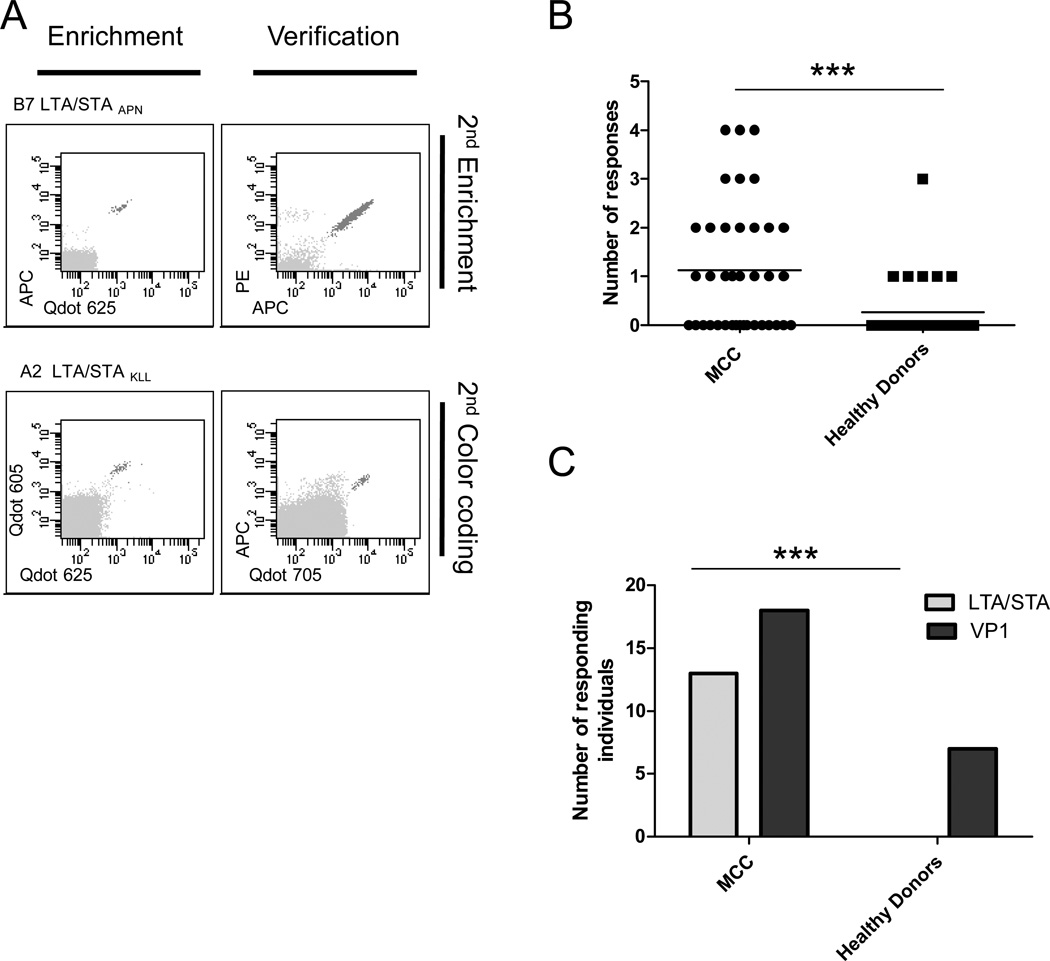
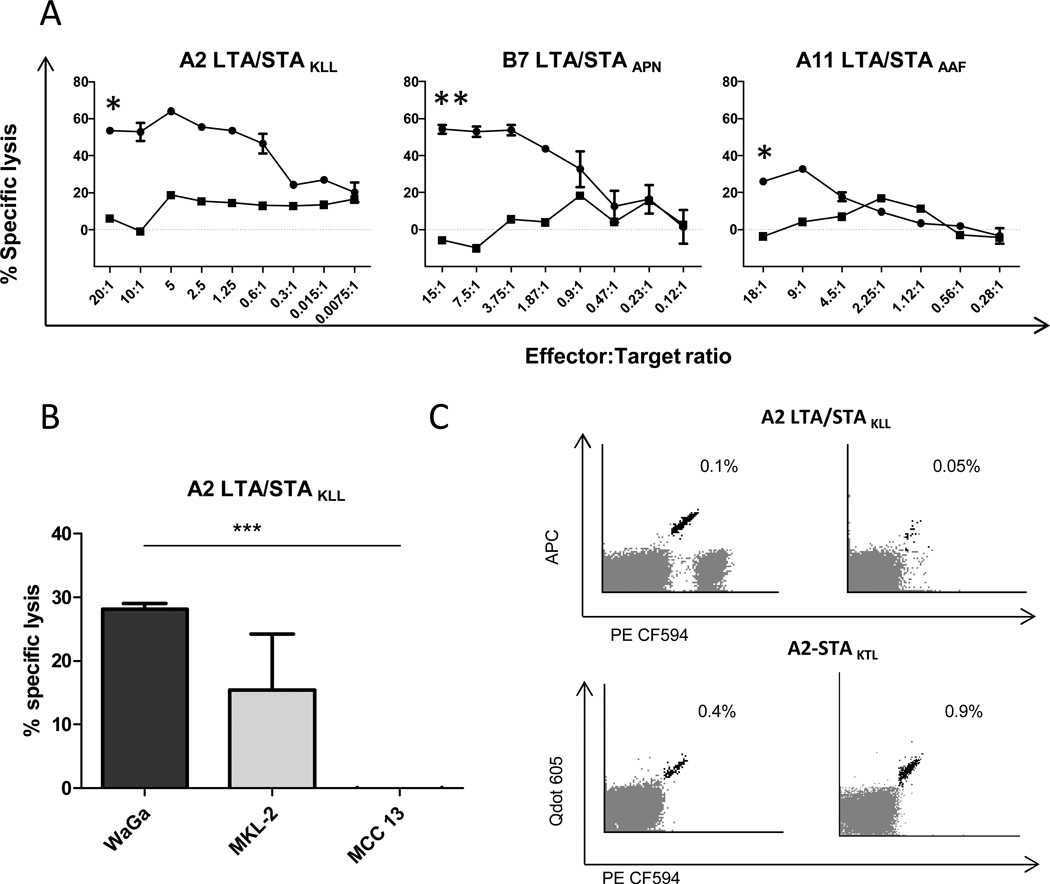
Results: In peripheral blood from 38 patients with MCC and 30 healthy donors, we identified 53 MCPyV-directed CD8 T-cell responses against 35 different peptide sequences. Strikingly, T-cell responses against oncoproteins were exclusively present in patients with MCC, but not in healthy donors. We further demonstrate both the processing and presentation of the oncoprotein-derived epitopes, as well as the lytic activity of oncoprotein-specific T cells toward MHC-matched MCC cells. Demonstrating the presence of oncoprotein-specific T cells among tumor-infiltrating lymphocytes further substantiated the relevance of the identified epitopes.
Conclusion: These T-cell epitopes represent ideal targets for antigen-specific immune therapy of MCC, and enable tracking and characterization of MCPyV-specific immune responses.
©2014 AACR.
Conflict of interest statement
The authors declare no conflicts of interest
Figures

References
-
- Friedlaender MM, Rubinger D, Rosenbaum E, Amir G, Siguencia E. Temporary regression of Merkel cell carcinoma metastases after cessation of cyclosporine. Transplantation. 2002;73:1849–1850. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
