

Wnt secretion from epithelial cells and subepithelial myofibroblasts is not required in the mouse intestinal stem cell niche in vivo
- PMID: 24527386
- PMCID: PMC3923227
- DOI: 10.1016/j.stemcr.2013.12.012
Wnt secretion from epithelial cells and subepithelial myofibroblasts is not required in the mouse intestinal stem cell niche in vivo
Abstract
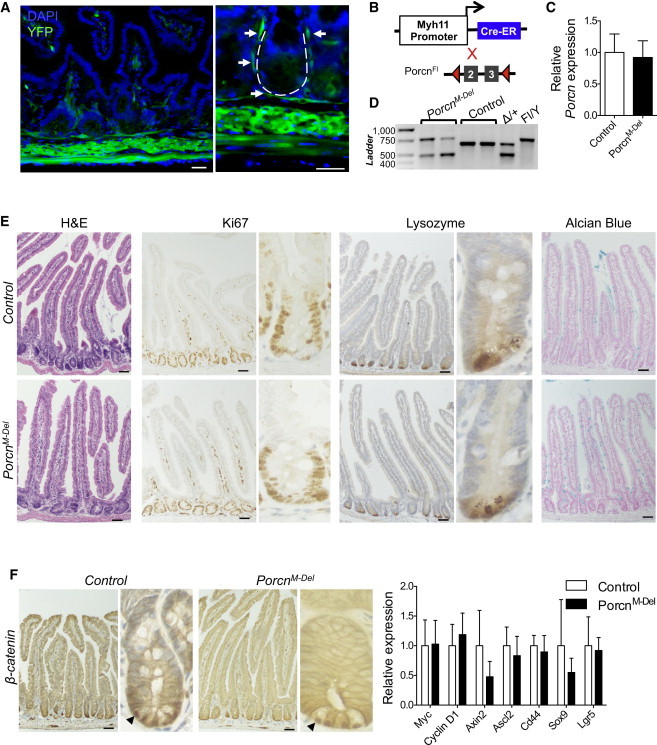
Wnt signaling is a crucial aspect of the intestinal stem cell niche required for crypt cell proliferation and differentiation. Paneth cells or subepithelial myofibroblasts are leading candidate sources of the required Wnt ligands, but this has not been tested in vivo. To abolish Wnt-ligand secretion, we used Porcupine (Porcn) conditional-null mice crossed to strains expressing inducible Cre recombinase in the epithelium, including Paneth cells (Villin-Cre (ERT2) ); in smooth muscle, including subepithelial myofibroblasts (Myh11-Cre (ERT2) ); and simultaneously in both compartments. Elimination of Wnt secretion from any of these compartments did not disrupt tissue morphology, cell proliferation, differentiation, or Wnt pathway activity. Thus, Wnt-ligand secretion from these cell populations is dispensable for intestinal homeostasis, revealing that a minor cell type or significant and unexpected redundancy is responsible for physiologic Wnt signaling in vivo.
Figures



References
-
- Barker N., van Es J.H., Kuipers J., Kujala P., van den Born M., Cozijnsen M., Haegebarth A., Korving J., Begthel H., Peters P.J., Clevers H. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature. 2007;449:1003–1007. - PubMed
-
- Barker N., Ridgway R.A., van Es J.H., van de Wetering M., Begthel H., van den Born M., Danenberg E., Clarke A.R., Sansom O.J., Clevers H. Crypt stem cells as the cells-of-origin of intestinal cancer. Nature. 2009;457:608–611. - PubMed
-
- Biechele S., Cox B.J., Rossant J. Porcupine homolog is required for canonical Wnt signaling and gastrulation in mouse embryos. Dev. Biol. 2011;355:275–285. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
