

A hydrolase of trehalose dimycolate induces nutrient influx and stress sensitivity to balance intracellular growth of Mycobacterium tuberculosis
- PMID: 24528862
- PMCID: PMC3974621
- DOI: 10.1016/j.chom.2014.01.008
A hydrolase of trehalose dimycolate induces nutrient influx and stress sensitivity to balance intracellular growth of Mycobacterium tuberculosis
Abstract
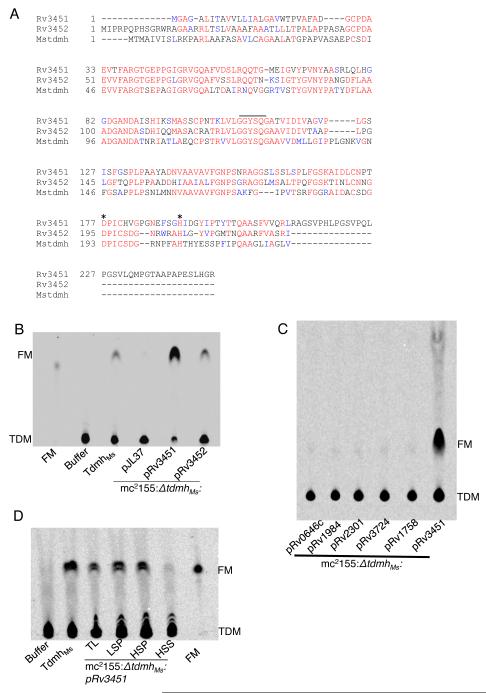
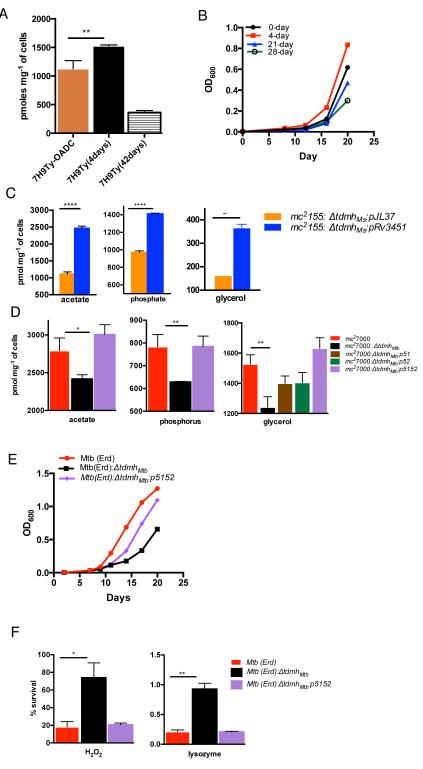
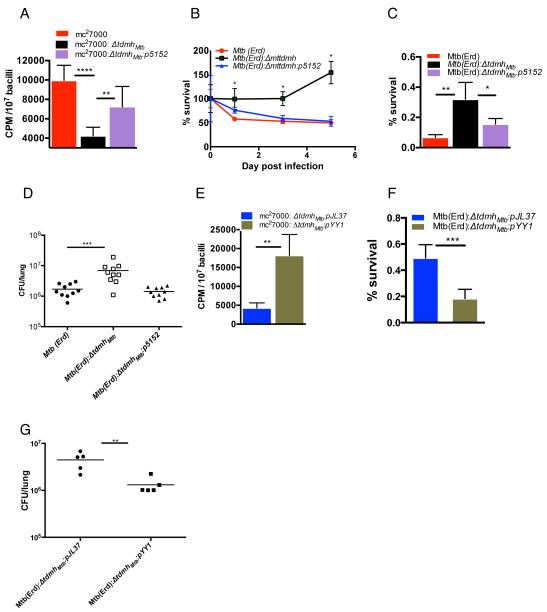
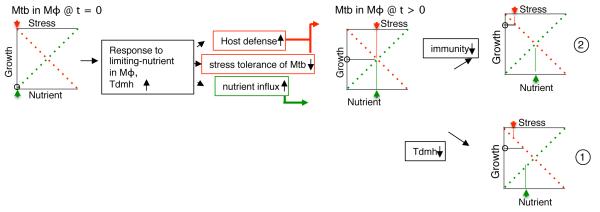
Chronic tuberculosis in an immunocompetent host is a consequence of the delicately balanced growth of Mycobacterium tuberculosis (Mtb) in the face of host defense mechanisms. We identify an Mtb enzyme (TdmhMtb) that hydrolyzes the mycobacterial glycolipid trehalose dimycolate and plays a critical role in balancing the intracellular growth of the pathogen. TdmhMtb is induced under nutrient-limiting conditions and remodels the Mtb envelope to increase nutrient influx but concomitantly sensitizes Mtb to stresses encountered in the host. Consistent with this, a ΔtdmhMtb mutant is more resilient to stress and grows to levels higher than those of wild-type in immunocompetent mice. By contrast, mutant growth is retarded in MyD88(-/-) mice, indicating that TdmhMtb provides a growth advantage to intracellular Mtb in an immunocompromised host. Thus, the effects and countereffects of TdmhMtb play an important role in balancing intracellular growth of Mtb in a manner that is directly responsive to host innate immunity.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures



References
-
- Akoh CC, Lee GC, Liaw YC, Huang TH, Shaw JF. GDSL family of serine esterases/lipases. Prog Lipid Res. 2004;43:534–552. - PubMed
-
- Bardarov S, Bardarov S, Jr., Pavelka MS, Jr., Sambandamurthy V, Larsen M, Tufariello J, Chan J, Hatfull G, Jacobs WR., Jr. Specialized transduction: an efficient method for generating marked and unmarked targeted gene disruptions in Mycobacterium tuberculosis, M. bovis BCG and M. smegmatis. Microbiology. 2002;148:3007–3017. - PubMed
-
- Belisle JT, Vissa VD, Sievert T, Takayama K, Brennan PJ, Besra GS. Role of the major antigen of Mycobacterium tuberculosis in cell wall biogenesis. Science. 1997;276:1420–1422. - PubMed
-
- Betts JC, Lukey PT, Robb LC, McAdam RA, Duncan K. Evaluation of a nutrient starvation model of Mycobacterium tuberculosis persistence by gene and protein expression profiling. Mol Microbiol. 2002;43:717–731. - PubMed
-
- Beutler B. Innate immunity: an overview. Mol Immunol. 2004;40:845–859. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
