

Measurement of lipolysis
- PMID: 24529439
- PMCID: PMC4018506
- DOI: 10.1016/B978-0-12-800280-3.00010-4
Measurement of lipolysis
Abstract
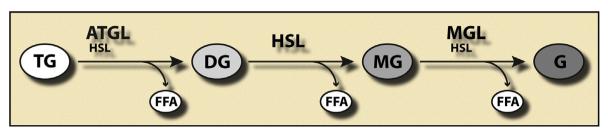
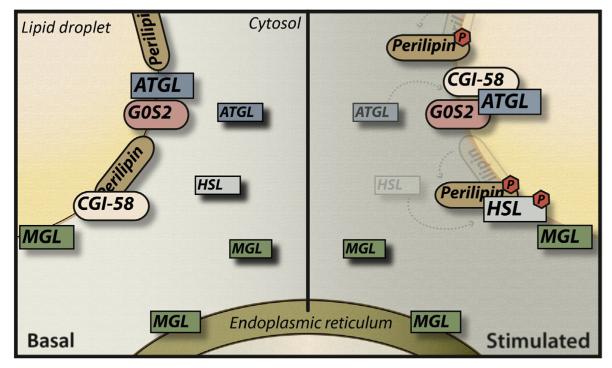
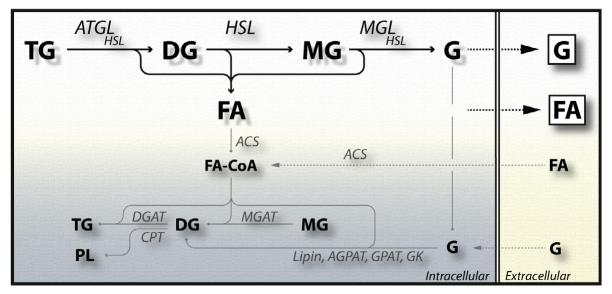
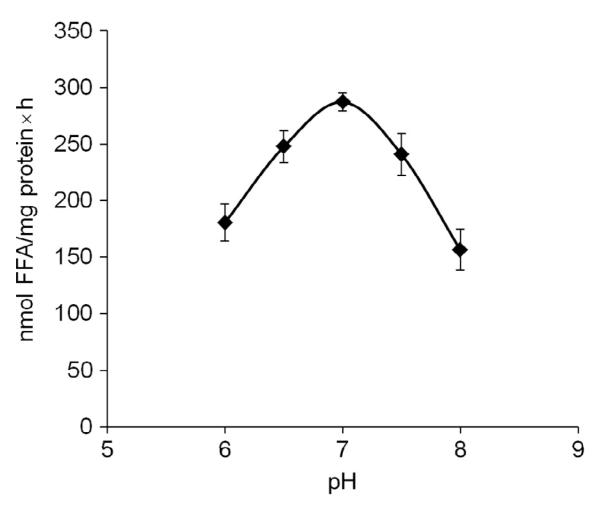
Lipolysis is defined as the hydrolytic cleavage of ester bonds in triglycerides (TGs), resulting in the generation of fatty acids (FAs) and glycerol. The two major TG pools in the body of vertebrates comprise intracellular TGs and plasma/nutritional TGs. Accordingly, this leads to the discrimination between intracellular and intravascular/gastrointestinal lipolysis, respectively. This chapter focuses exclusively on intracellular lipolysis, referred to as lipolysis herein. The lipolytic cleavage of TGs occurs in essentially all cells and tissues of the body. In all of them, the resulting FAs are utilized endogenously for energy production or biosynthetic pathways with one exception, white adipose tissue (WAT). WAT releases FAs and glycerol to supply nonadipose tissues at times of nutrient deprivation. The fundamental role of lipolysis in lipid and energy homeostasis requires the accurate measurement of lipase activities and lipolytic rates. The recent discovery of new enzymes and regulators that mediate the hydrolysis of TG has made these measurements more complex. Here, we describe detailed methodology for how to measure lipolysis and specific enzymes' activities in cells, organs, and their respective extracts.
Keywords: ATGL; Adipose tissue; HSL; Lipase; Lipid droplet; Lipolysis; MGL; Triglyceride hydrolase activity.
© 2014 Elsevier Inc. All rights reserved.
Figures
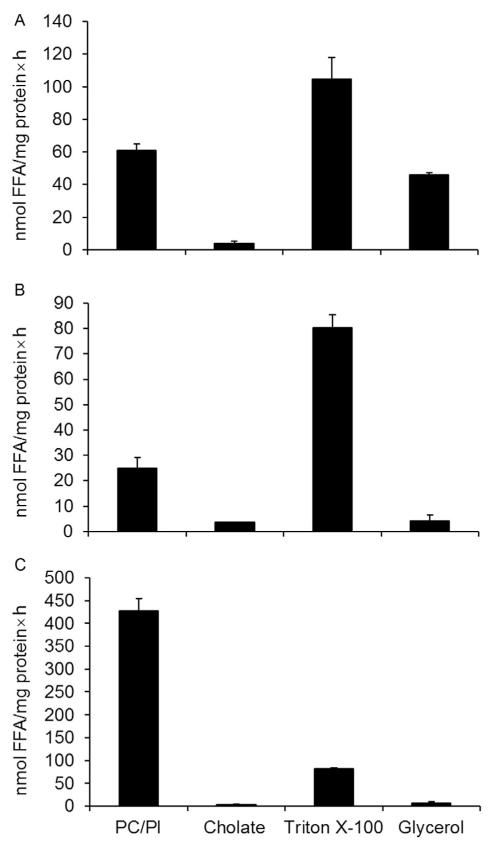
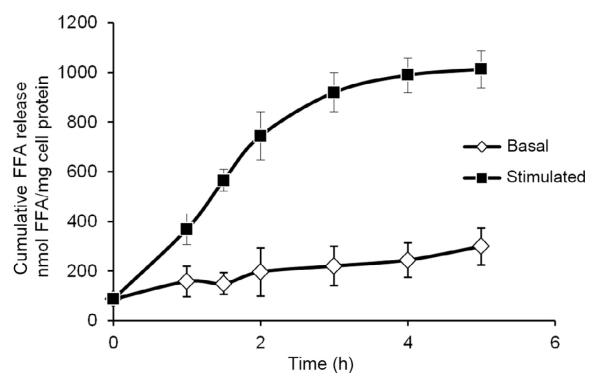
References
-
- Clifford GM, Londos C, Kraemer FB, Vernon RG, Yeaman SJ. Translocation of hormone-sensitive lipase and perilipin upon lipolytic stimulation of rat adipocytes. The Journal of Biological Chemistry. 2000;275:5011–5015. - PubMed
-
- Ebdrup S, Sørensen LG, Olsen OH, Jacobsen P. Synthesis and structure-activity relationship for a novel class of potent and selective carbamoyl-triazole based inhibitors of hormone sensitive lipase. Journal of Medicinal Chemistry. 2004;47:400–410. - PubMed
-
- Egan JJ, Greenberg AS, Chang MK, Wek SA, Moos MCJ, Londos C. Mechanism of hormone-stimulated lipolysis in adipocytes: Translocation of hormone-sensitive lipase to the lipid storage droplet. Proceedings of the National Academy of Sciences of the United States of America. 1992;89:8537–8541. - PMC - PubMed
-
- Granneman JG, Moore HP, Granneman RL, Greenberg AS, Obin MS, Zhu Z. Analysis of lipolytic protein trafficking and interactions in adipocytes. Journal of Biological Chemistry. 2006;282:5726–5735. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
