

Effects of modulators of AMP-activated protein kinase on TASK-1/3 and intracellular Ca(2+) concentration in rat carotid body glomus cells
- PMID: 24530802
- PMCID: PMC3998119
- DOI: 10.1016/j.resp.2014.01.020
Effects of modulators of AMP-activated protein kinase on TASK-1/3 and intracellular Ca(2+) concentration in rat carotid body glomus cells
Abstract
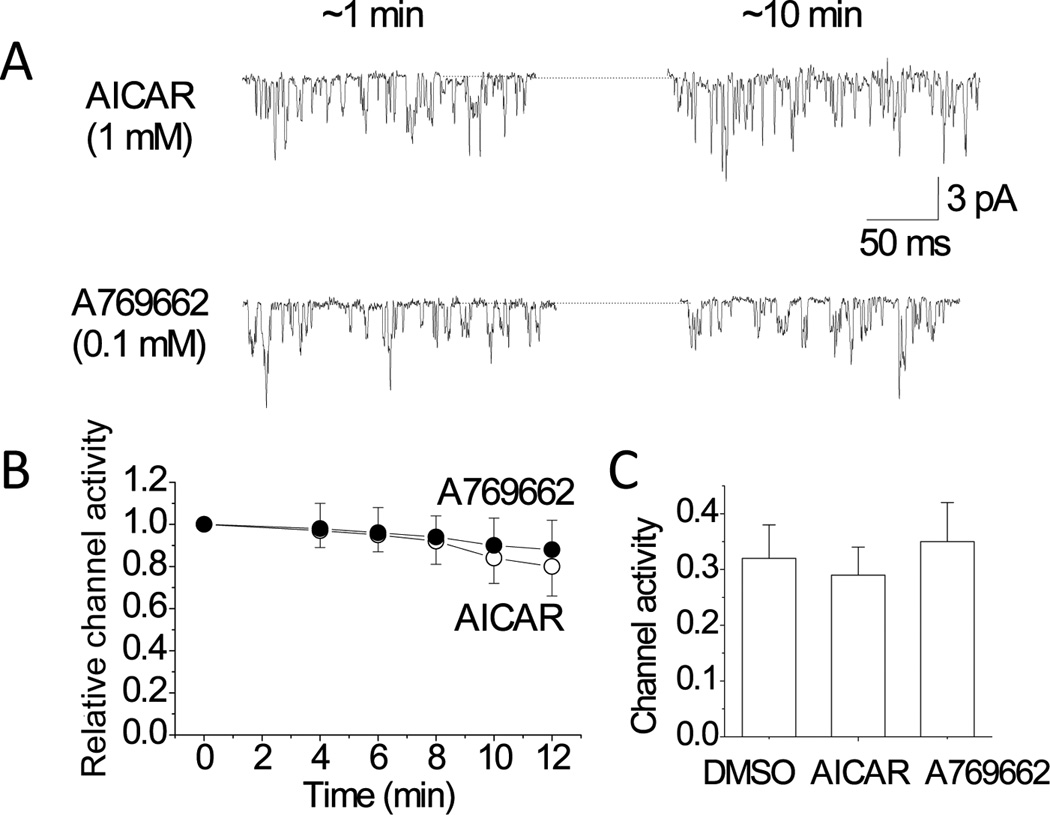
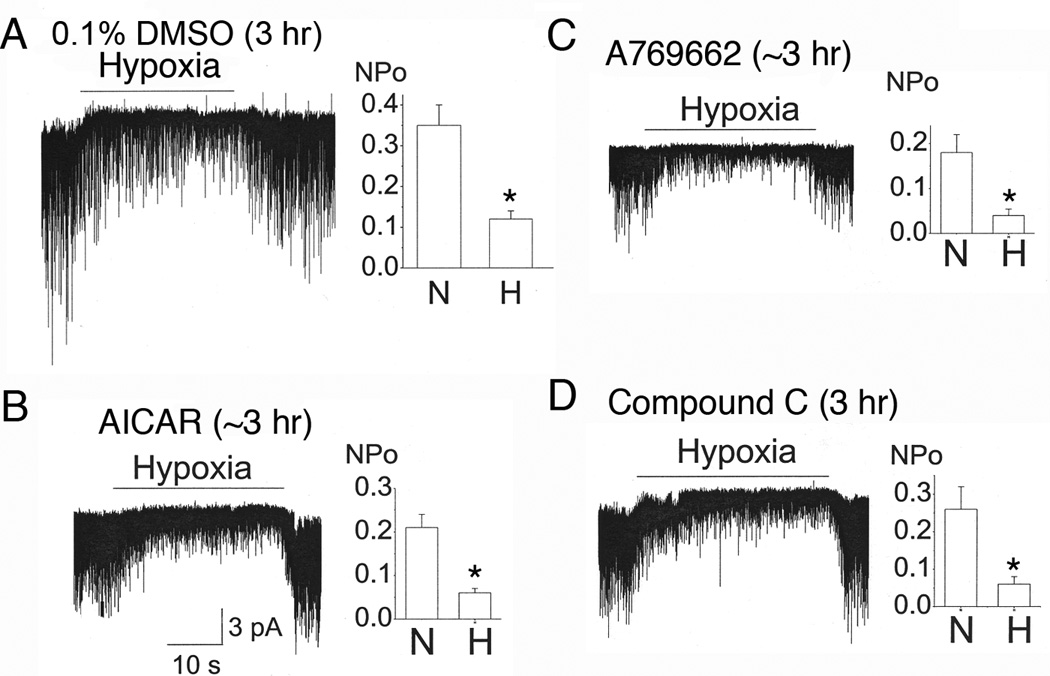
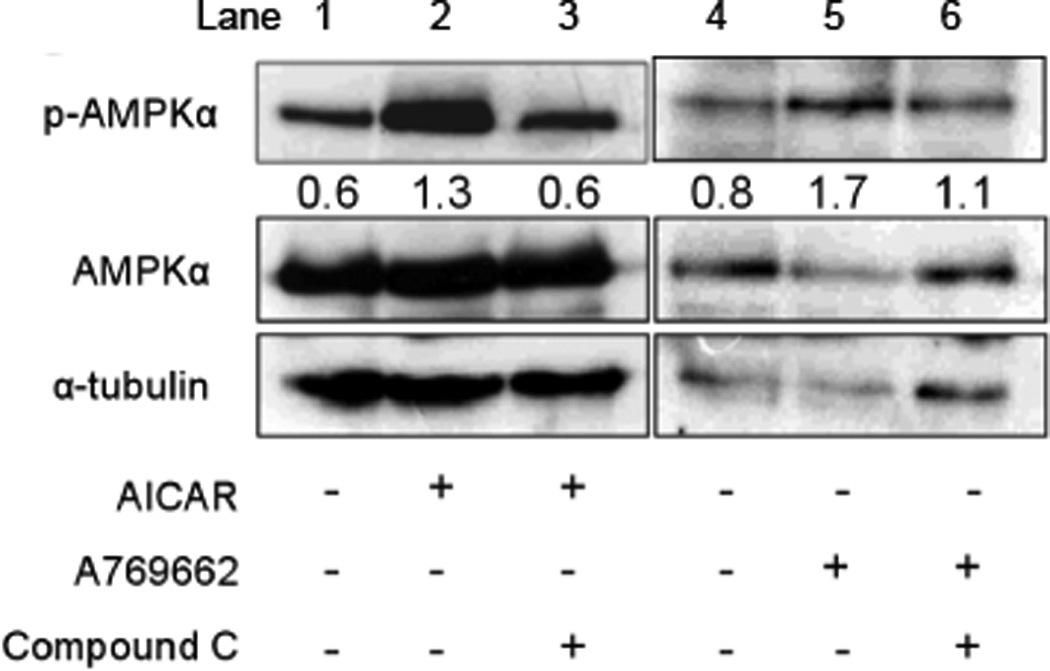
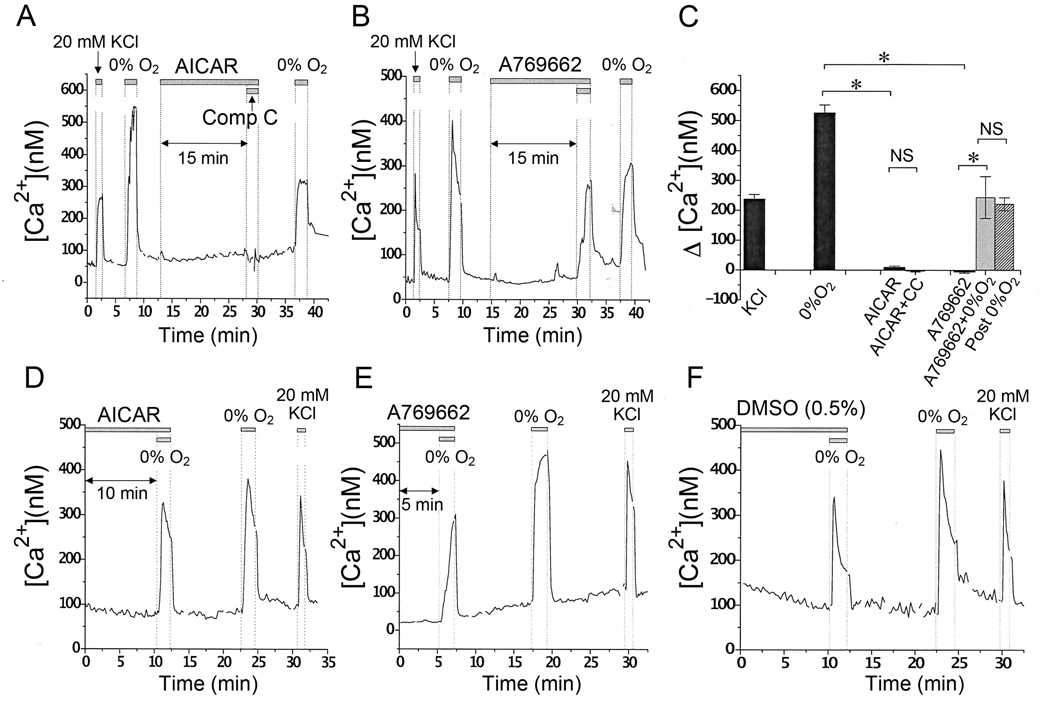
Acute hypoxia depolarizes carotid body chemoreceptor (glomus) cells and elevates intracellular Ca(2+) concentration ([Ca(2+)]i). Recent studies suggest that AMP-activated protein kinase (AMPK) mediates these effects of hypoxia by inhibiting the background K(+) channels such as TASK. Here we studied the effects of modulators of AMPK on TASK activity in cell-attached patches. Activators of AMPK (1mM AICAR and 0.1-0.5mM A769662) did not inhibit TASK activity or cause depolarization during acute (10min) or prolonged (2-3h) exposure. Hypoxia inhibited TASK activity by ∼70% in cells pretreated with AICAR or A769662. Both AICAR and A769662 (15-40min) failed to increase [Ca(2+)]i in glomus cells. Compound C (40μM), an inhibitor of AMPK, showed no effect on hypoxia-induced inhibition of TASK. AICAR and A769662 phosphorylated AMPKα in PC12 cells, and Compound C blocked the phosphorylation. Our results suggest that AMPK does not affect TASK activity and is not involved in hypoxia-induced elevation of intracellular [Ca(2+)] in isolated rat carotid body glomus cells.
Keywords: AMP kinase; Background K(+) channels; Carotid body; Chemoreceptors; Hypoxia.
Copyright © 2014 Elsevier B.V. All rights reserved.
Figures
References
-
- Buckler KJ. TASK-like potassium channels and oxygen sensing in the carotid body. Respir Physiol Neurobiol. 2007;157:55–64. - PubMed
-
- Corton JM, Gillespie JG, Hawley SA, Hardie DG. 5-aminoimidazole-4-carboxamide ribonucleoside. A specific method for activating AMP-activated protein kinase in intact cells? Eur J Biochem. 1995;229:558–565. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
