

A temporal basis for predicting the sensory consequences of motor commands in an electric fish
- PMID: 24531306
- PMCID: PMC4070001
- DOI: 10.1038/nn.3650
A temporal basis for predicting the sensory consequences of motor commands in an electric fish
Abstract
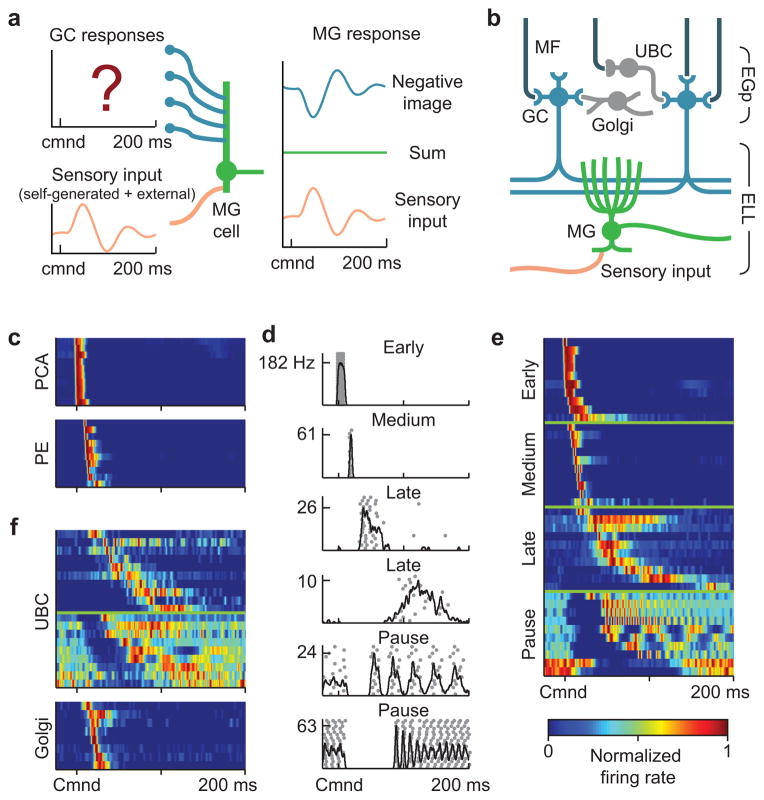
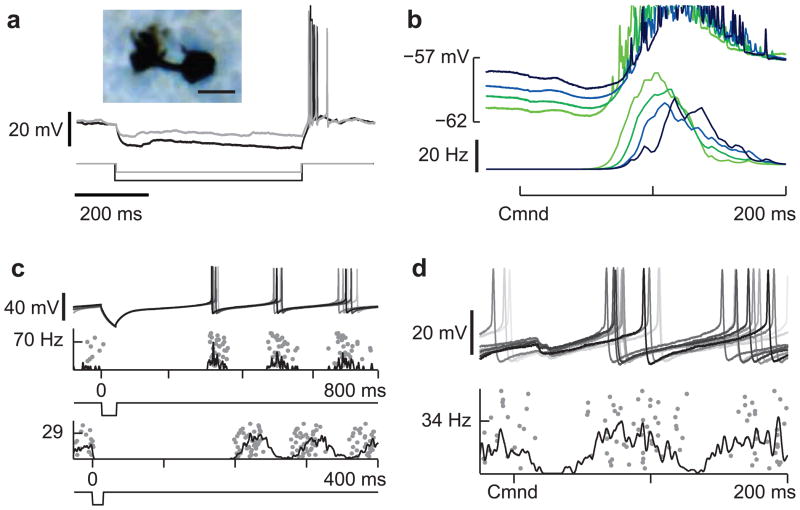
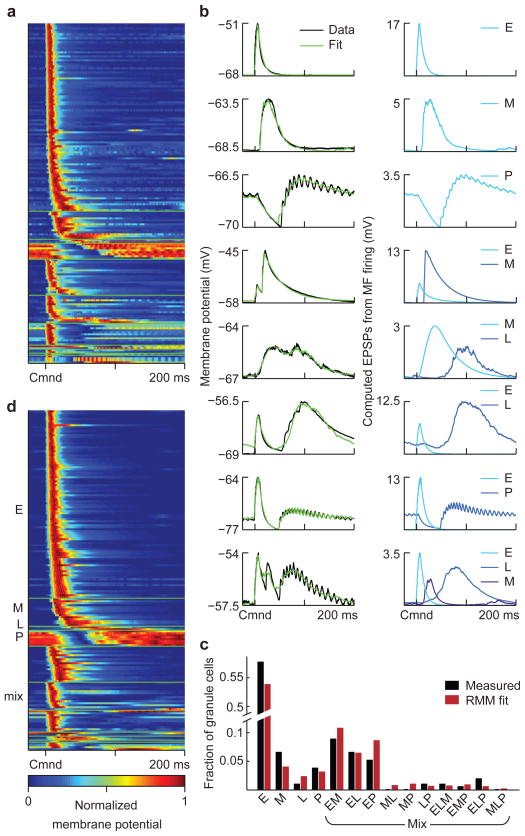
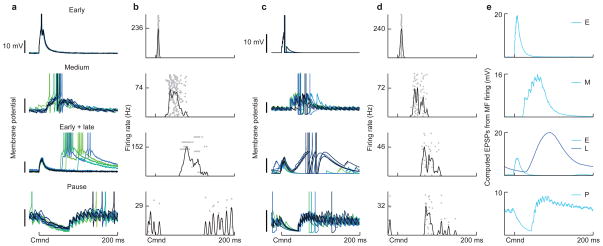
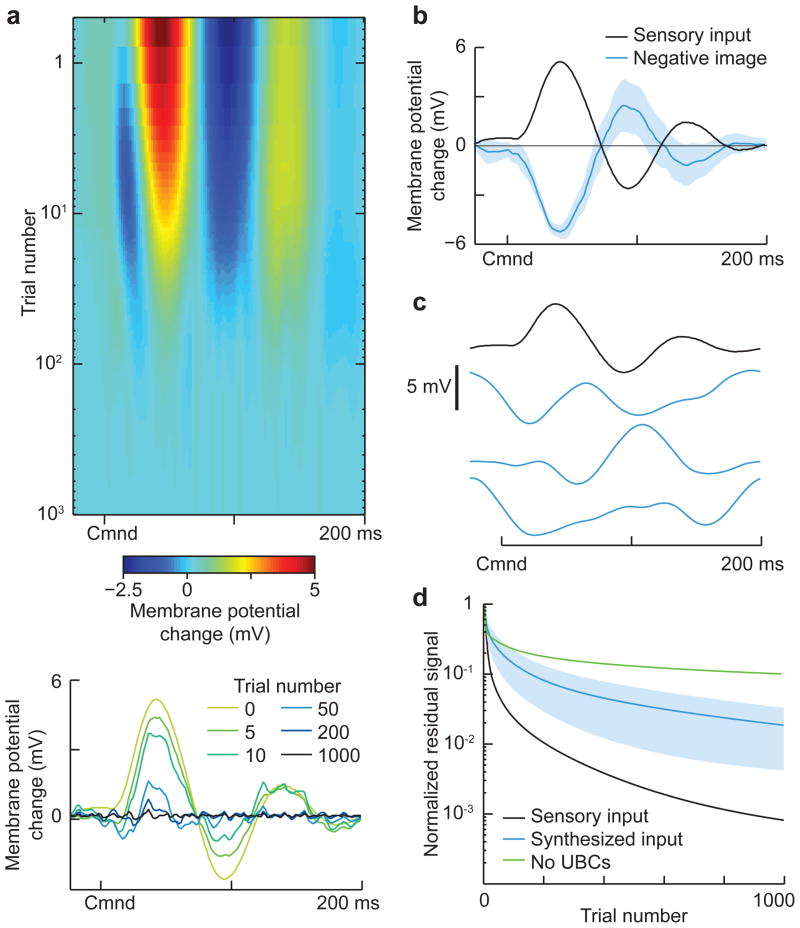
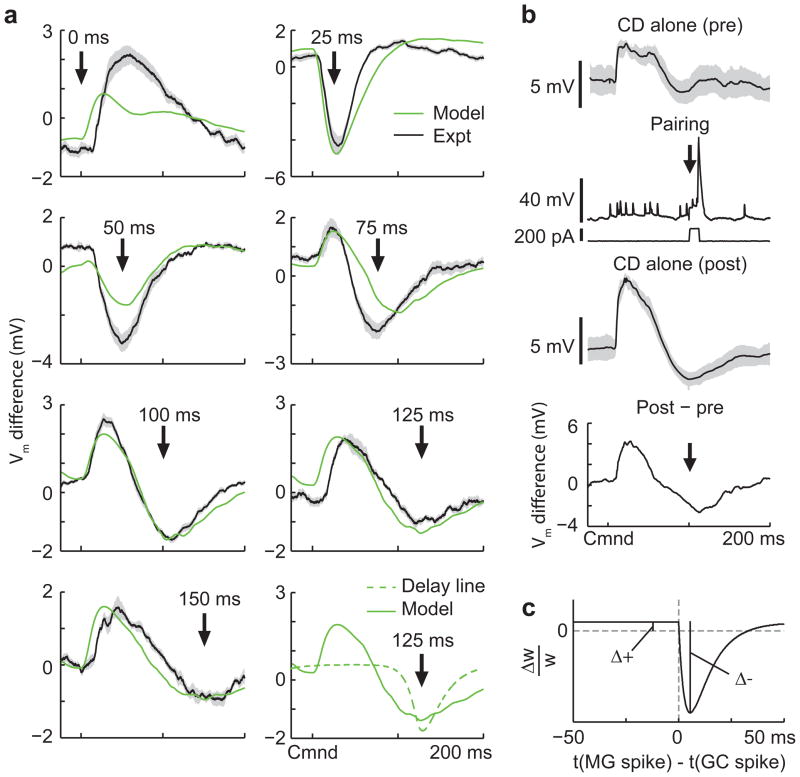
Mormyrid electric fish are a model system for understanding how neural circuits predict the sensory consequences of motor acts. Medium ganglion cells in the electrosensory lobe create negative images that predict sensory input resulting from the fish's electric organ discharge (EOD). Previous studies have shown that negative images can be created through plasticity at granule cell-medium ganglion cell synapses, provided that granule cell responses to the brief EOD command are sufficiently varied and prolonged. Here we show that granule cells indeed provide such a temporal basis and that it is well-matched to the temporal structure of self-generated sensory inputs, allowing rapid and accurate sensory cancellation and explaining paradoxical features of negative images. We also demonstrate an unexpected and critical role of unipolar brush cells (UBCs) in generating the required delayed responses. These results provide a mechanistic account of how copies of motor commands are transformed into sensory predictions.
Figures
Similar articles
-
Signal Diversification Is Associated with Corollary Discharge Evolution in Weakly Electric Fish.J Neurosci. 2020 Aug 12;40(33):6345-6356. doi: 10.1523/JNEUROSCI.0875-20.2020. Epub 2020 Jul 13. J Neurosci. 2020. PMID: 32661026 Free PMC article.
-
Sensory processing and corollary discharge effects in the mormyromast regions of the mormyrid electrosensory lobe. I. Field potentials, cellular activity in associated structures.J Neurophysiol. 1992 Sep;68(3):843-58. doi: 10.1152/jn.1992.68.3.843. J Neurophysiol. 1992. PMID: 1432052
-
Sensory processing and corollary discharge effects in mormyromast regions of mormyrid electrosensory lobe. II. Cell types and corollary discharge plasticity.J Neurophysiol. 1992 Sep;68(3):859-75. doi: 10.1152/jn.1992.68.3.859. J Neurophysiol. 1992. PMID: 1432053
-
Synaptic plasticity in the mormyrid electrosensory lobe.J Exp Biol. 1999 May;202(Pt 10):1339-47. doi: 10.1242/jeb.202.10.1339. J Exp Biol. 1999. PMID: 10210674 Review.
-
The role of motor command feedback in electrosensory processing.Eur J Morphol. 1994 Aug;32(2-4):225-34. Eur J Morphol. 1994. PMID: 7803171 Review.
Cited by
-
ON and OFF unipolar brush cells transform multisensory inputs to the auditory system.Neuron. 2015 Mar 4;85(5):1029-42. doi: 10.1016/j.neuron.2015.02.009. Neuron. 2015. PMID: 25741727 Free PMC article.
-
A History of Corollary Discharge: Contributions of Mormyrid Weakly Electric Fish.Front Integr Neurosci. 2020 Jul 29;14:42. doi: 10.3389/fnint.2020.00042. eCollection 2020. Front Integr Neurosci. 2020. PMID: 32848649 Free PMC article. Review.
-
Constructing functional models from biophysically-detailed neurons.PLoS Comput Biol. 2022 Sep 8;18(9):e1010461. doi: 10.1371/journal.pcbi.1010461. eCollection 2022 Sep. PLoS Comput Biol. 2022. PMID: 36074765 Free PMC article.
-
Simple spike dynamics of Purkinje cells in the macaque vestibulo-cerebellum during passive whole-body self-motion.Proc Natl Acad Sci U S A. 2020 Feb 11;117(6):3232-3238. doi: 10.1073/pnas.1915873117. Epub 2020 Jan 27. Proc Natl Acad Sci U S A. 2020. PMID: 31988119 Free PMC article.
-
Intrinsic and Synaptic Properties Shaping Diverse Behaviors of Neural Dynamics.Front Comput Neurosci. 2020 Apr 21;14:26. doi: 10.3389/fncom.2020.00026. eCollection 2020. Front Comput Neurosci. 2020. PMID: 32372936 Free PMC article.
References
-
- Bell CC, Russell CJ. Effect of electric organ discharge on ampullary receptors in a mormyrid. Brain Res. 1978;145:85–96. - PubMed
-
- Bell CC. An efference copy modified by reafferent input. Science. 1981;214:450–453. - PubMed
-
- Roberts PD, Bell CC. Computational consequences of temporally asymmetric learning rules: II. sensory image cancellation. J Comput Neurosci. 2000;9:67–83. - PubMed
-
- Williams A, Roberts PD, Leen TK. Stability of negative-image equilibria in spike-timing-dependent plasticity. Phys Rev E Stat Nonlin Soft Matter Phys. 2003;68:021923. - PubMed
-
- Bell CC, Han VZ, Sugawara S, Grant K. Synaptic plasticity in a cerebellum-like structure depends on temporal order. Nature. 1997;387:278–281. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
