

Epimerization and substrate gating by a TE domain in β-lactam antibiotic biosynthesis
- PMID: 24531841
- PMCID: PMC3961552
- DOI: 10.1038/nchembio.1456
Epimerization and substrate gating by a TE domain in β-lactam antibiotic biosynthesis
Abstract
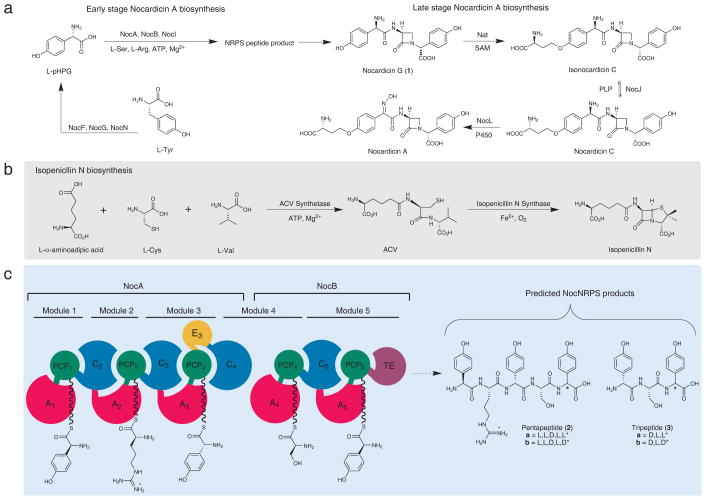
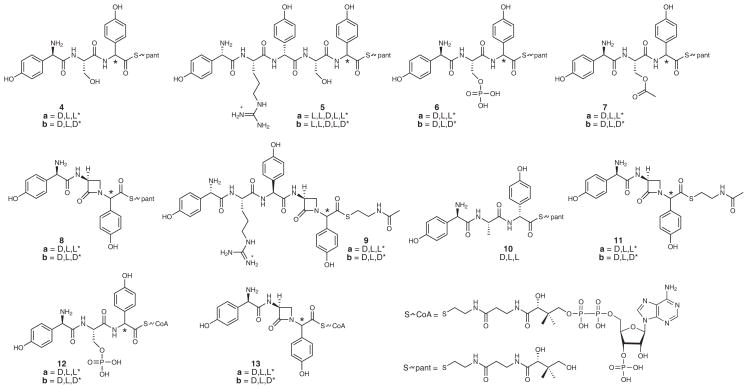
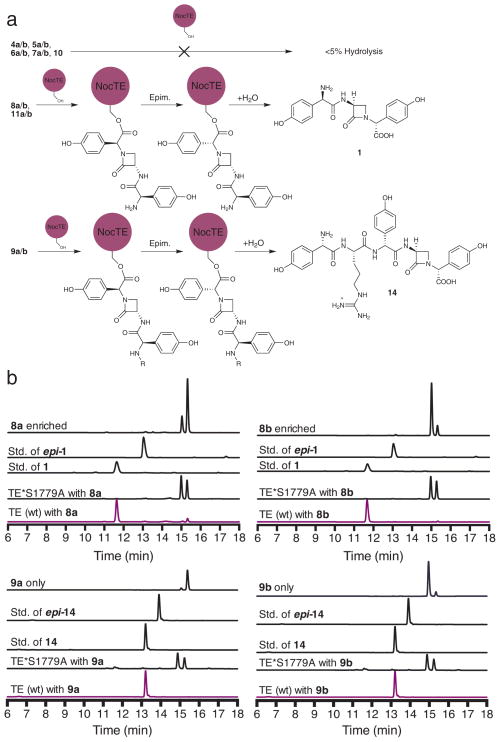
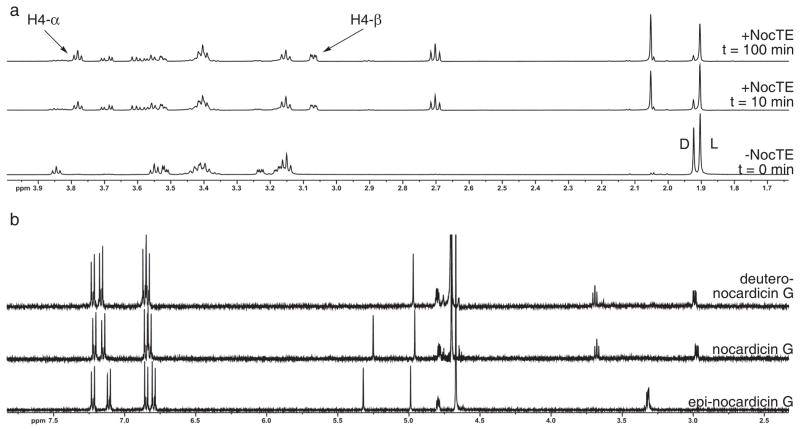
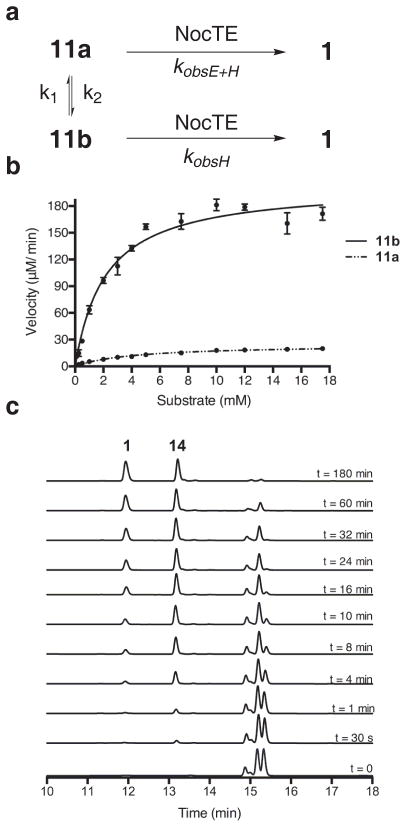
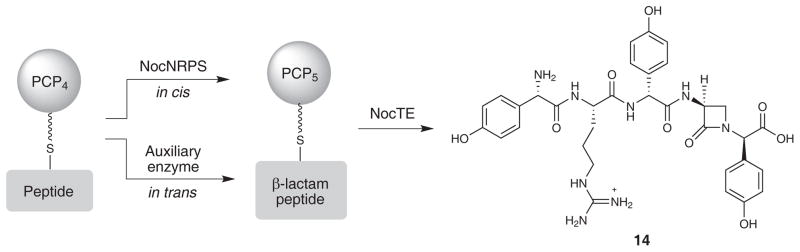
Nonribosomal peptide synthetases are versatile engines of bioactive natural product biosynthesis that function according to the multiple carrier thiotemplate mechanism. C-terminal thioesterase (TE) domains of these giant modular proteins typically catalyze product release by hydrolysis or macrocyclization. We now report an unprecedented, dual-function TE that is involved in the biosynthesis of nocardicin A, which is the paradigm monocyclic β-lactam antibiotic. Contrary to our expectation, a stereodefined series of potential peptide substrates for the nocardicin TE domain failed to undergo hydrolysis. The stringent discrimination against peptide intermediates was overcome by prior monocyclic β-lactam formation at an L-seryl site. Kinetic data are interpreted such that the TE domain acts as a gatekeeper to hold the assembling peptide on an upstream domain until β-lactam formation takes place and then rapidly catalyzes epimerization, which has not been observed previously as a TE catalytic function, and thioesterase cleavage to discharge a fully fledged pentapeptide β-lactam harboring nocardicin G, the universal precursor of the nocardicins.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Aoki H, Sakai H, Kohsaka M, Konomi T, Hosoda J. Nocardicin A, a New Monocyclic β-Lactam Antibiotic. I Discovery, Isolation and Characterization. J Antibiot. 1976;29:492–500. - PubMed
-
- Townsend CA, Wilson BA. The Role of Nocardicin G in Nocardicin A Biosynthesis. J Am Chem Soc. 1988;110:3320–3321.
-
- Townsend CA, Brown AM. Nocardicin A: Biosynthetic Experiments with Amino Acid Precursors. J Am Chem Soc. 1983;105:913–918.
-
- Townsend CA, Brown AM, Nguyen LT. Nocardicin A: Stereochemical and Biomimetic Studies of Monocyclic β-Lactam Formation. J Am Chem Soc. 1983;105:919–927.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
