

The transcription factor E4BP4 is not required for extramedullary pathways of NK cell development
- PMID: 24534532
- PMCID: PMC3948112
- DOI: 10.4049/jimmunol.1302765
The transcription factor E4BP4 is not required for extramedullary pathways of NK cell development
Abstract
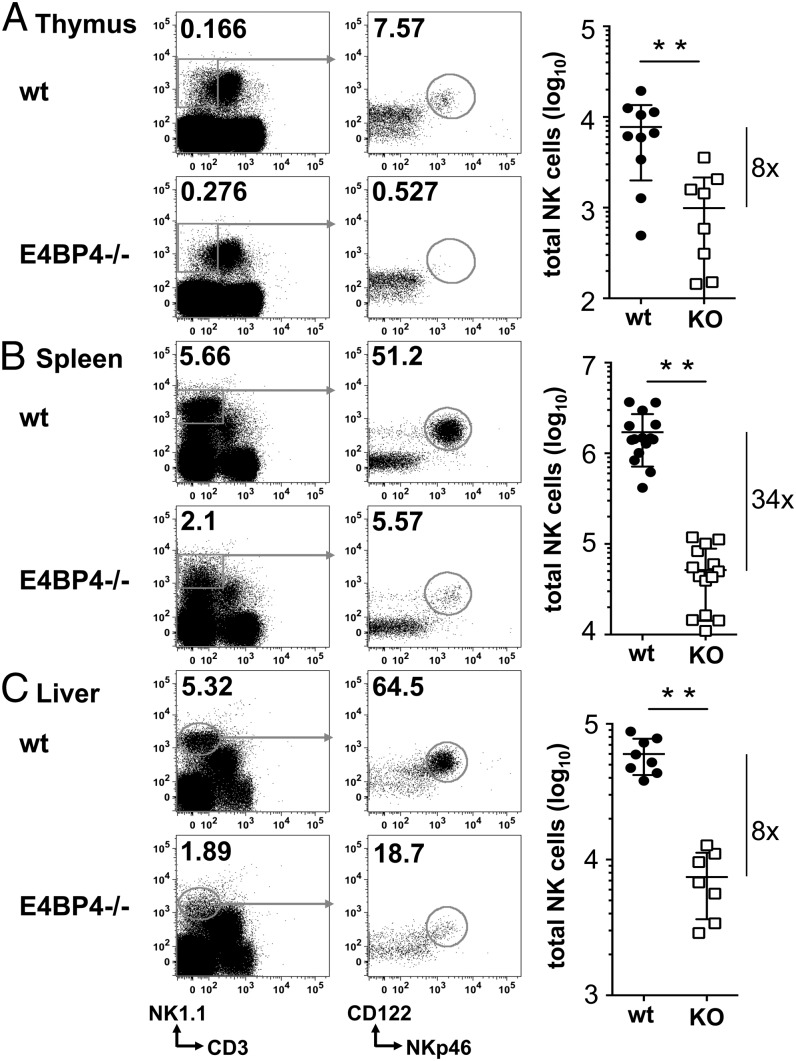
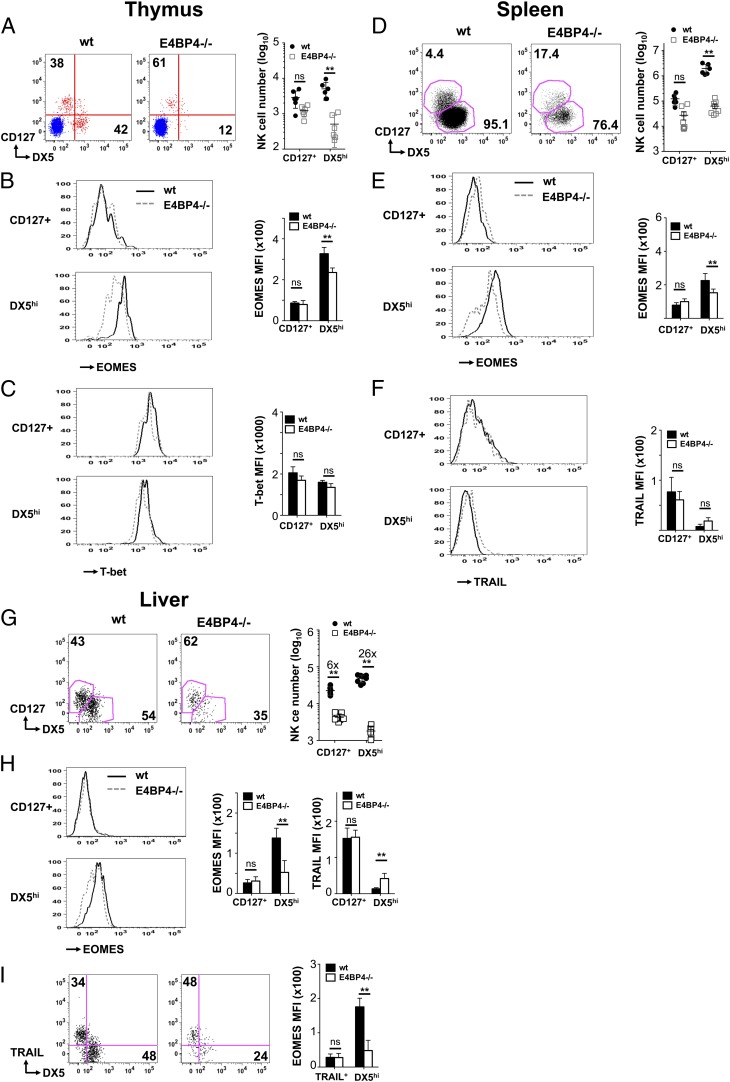
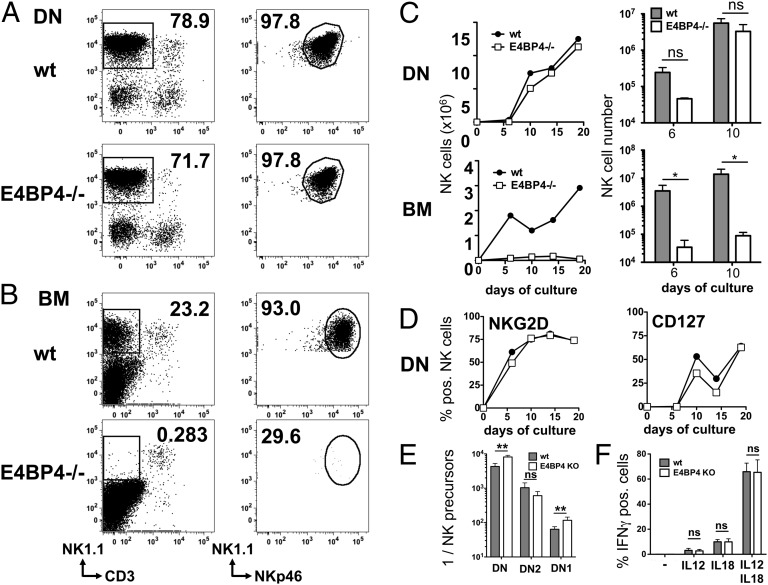
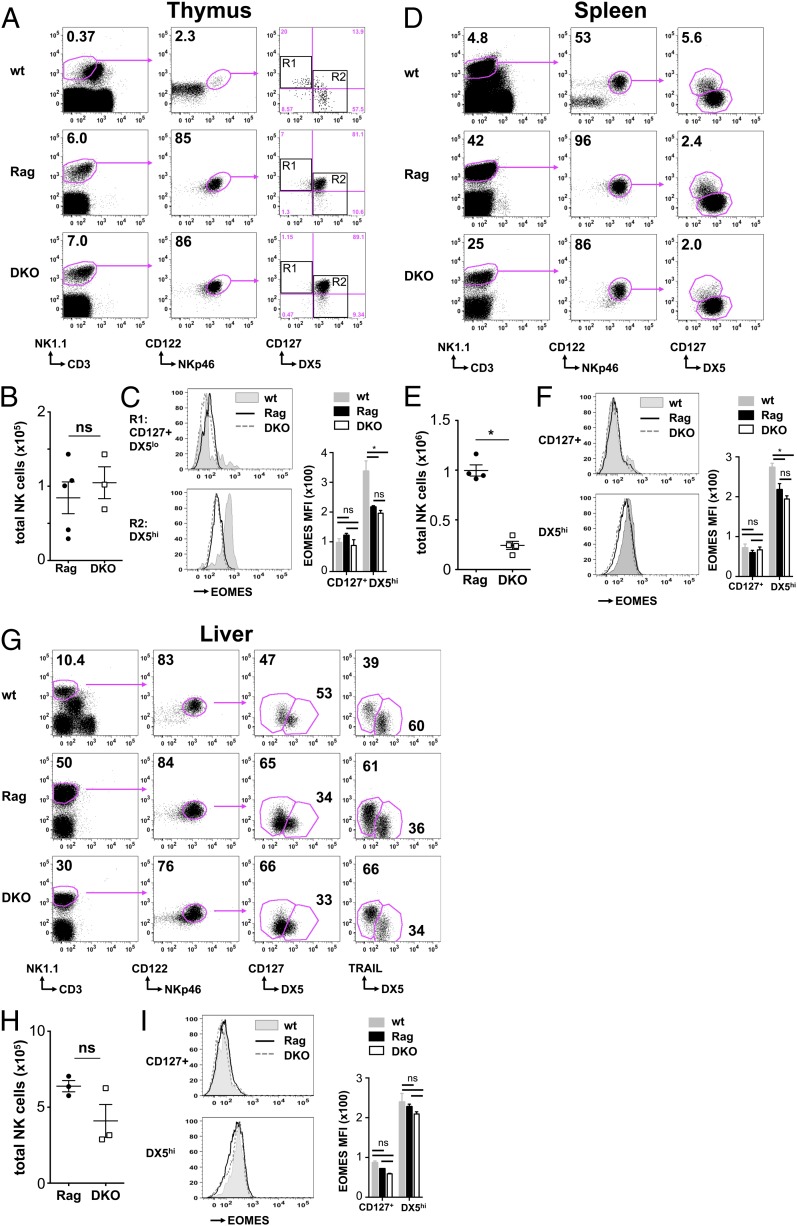
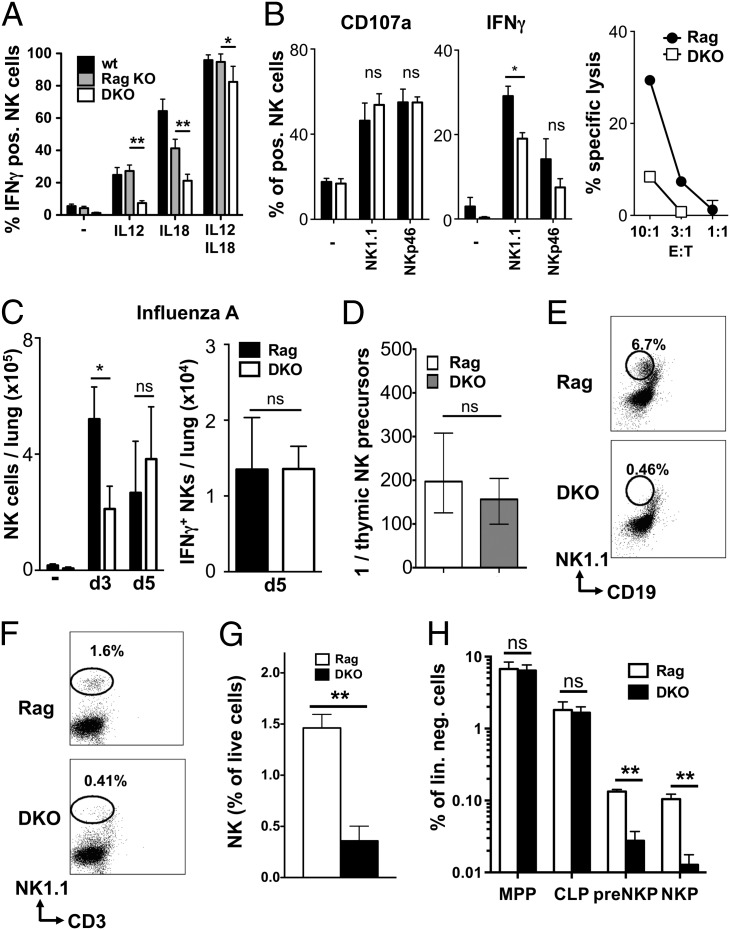
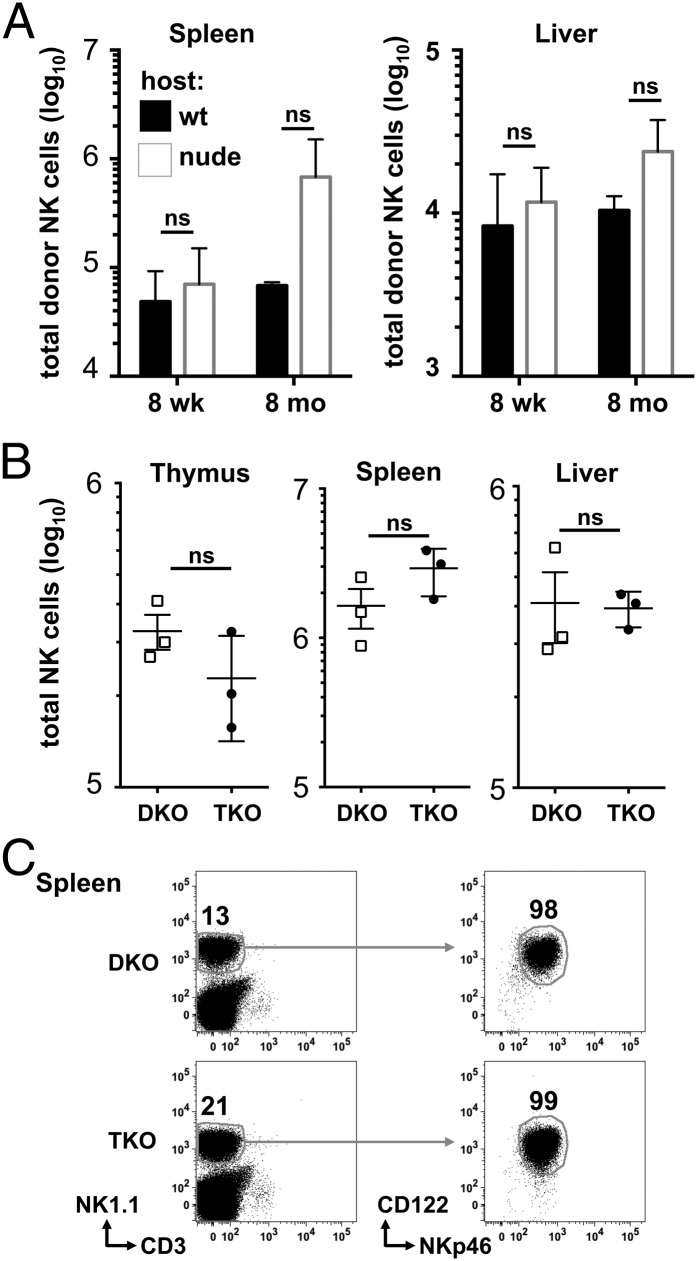
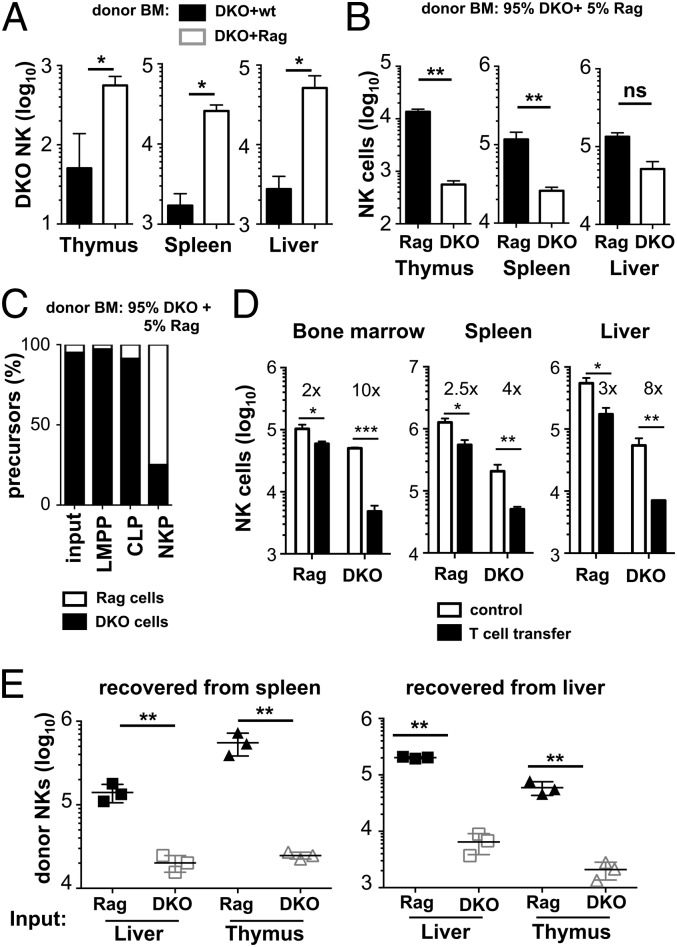
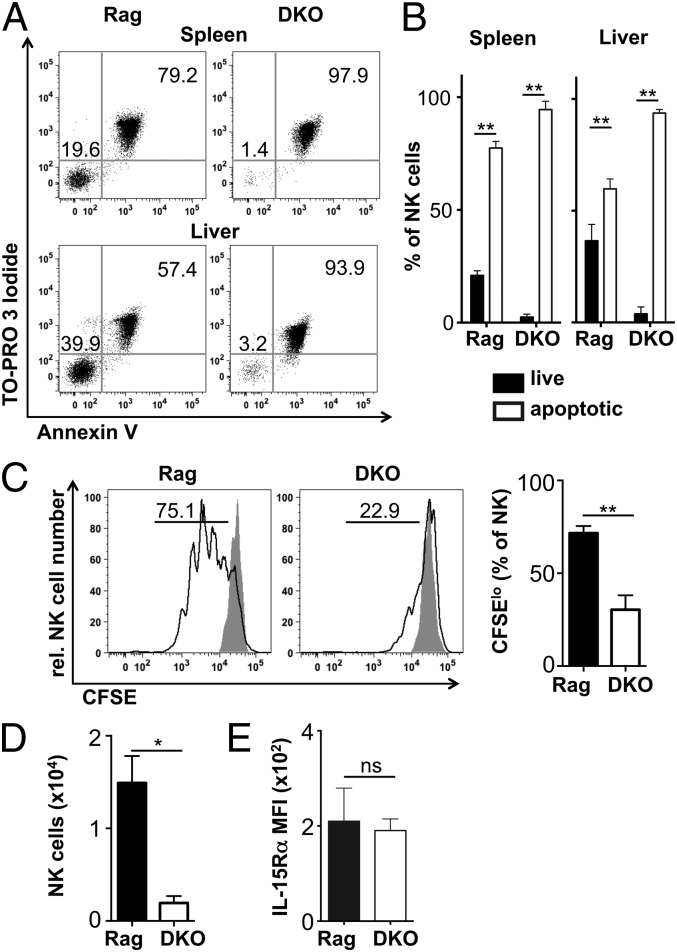
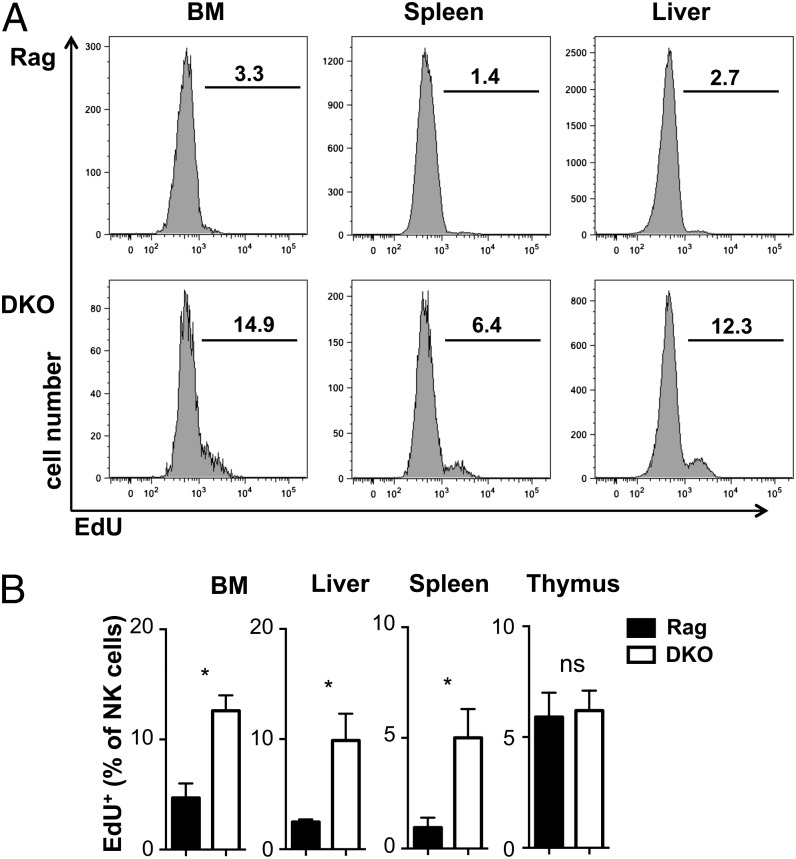
NK cells contribute to antitumor and antiviral immunosurveillance. Their development in the bone marrow (BM) requires the transcription factor E4BP4/NFIL3, but requirements in other organs are less well defined. In this study, we show that CD3(-)NK1.1(+)NKp46(+)CD122(+) NK cells of immature phenotype and expressing low eomesodermin levels are found in thymus, spleen, and liver of E4BP4-deficient mice, whereas numbers of mature, eomesodermin(high) conventional NK cells are drastically reduced. E4BP4-deficient CD44(+)CD25(-) double-negative 1 thymocytes efficiently develop in vitro into NK cells with kinetics, phenotype, and functionality similar to wild-type controls, whereas no NK cells develop from E4BP4-deficient BM precursors. In E4BP4/Rag-1 double-deficient (DKO) mice, NK cells resembling those in Rag-1-deficient controls are found in similar numbers in the thymus and liver. However, NK precursors are reduced in DKO BM, and no NK cells develop from DKO BM progenitors in vitro. DKO thymocyte precursors readily develop into NK cells, but DKO BM transfers into nude recipients and NK cells in E4BP4/Rag-1/IL-7 triple-KO mice indicated thymus-independent NK cell development. In the presence of T cells or E4BP4-sufficient NK cells, DKO NK cells have a selective disadvantage, and thymic and hepatic DKO NK cells show reduced survival when adoptively transferred into lymphopenic hosts. This correlates with higher apoptosis rates and lower responsiveness to IL-15 in vitro. In conclusion, we demonstrate E4BP4-independent development of NK cells of immature phenotype, reduced fitness, short t1/2, and potential extramedullary origin. Our data identify E4BP4-independent NK cell developmental pathways and a role for E4BP4 in NK cell homeostasis.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
