

Thyroid hormone signaling in vivo requires a balance between coactivators and corepressors
- PMID: 24550004
- PMCID: PMC3993596
- DOI: 10.1128/MCB.00129-14
Thyroid hormone signaling in vivo requires a balance between coactivators and corepressors
Abstract
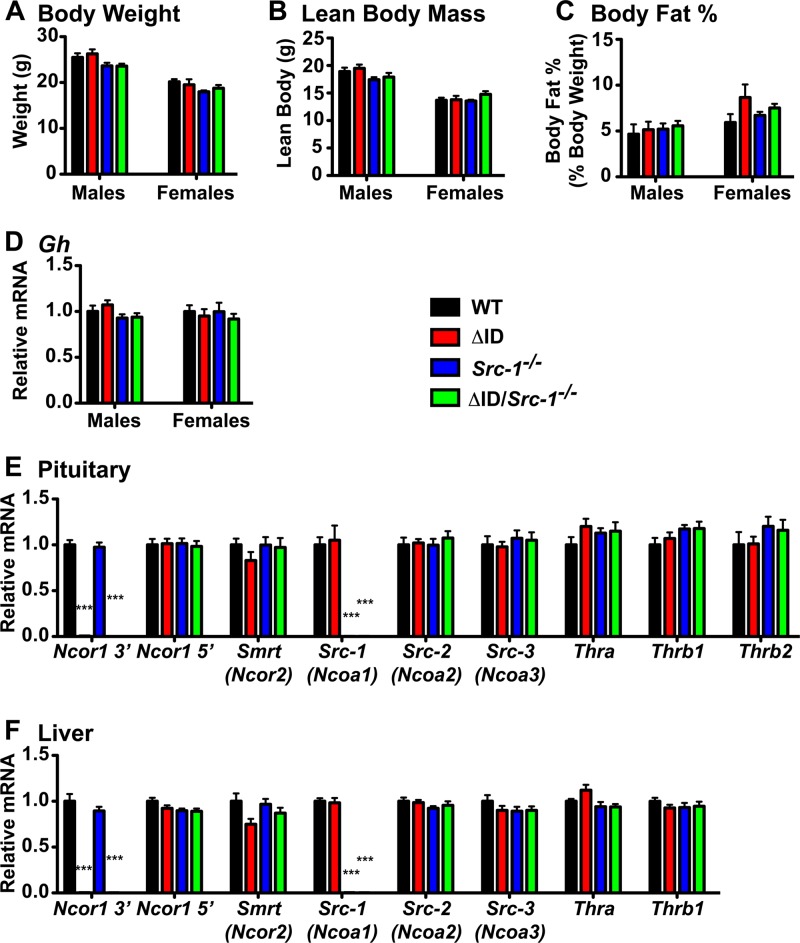
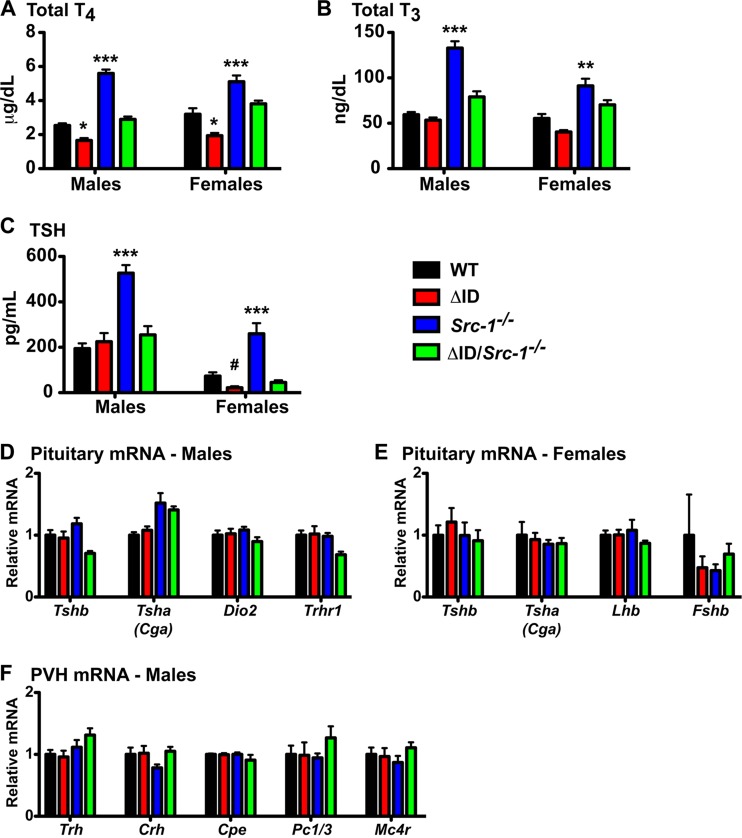
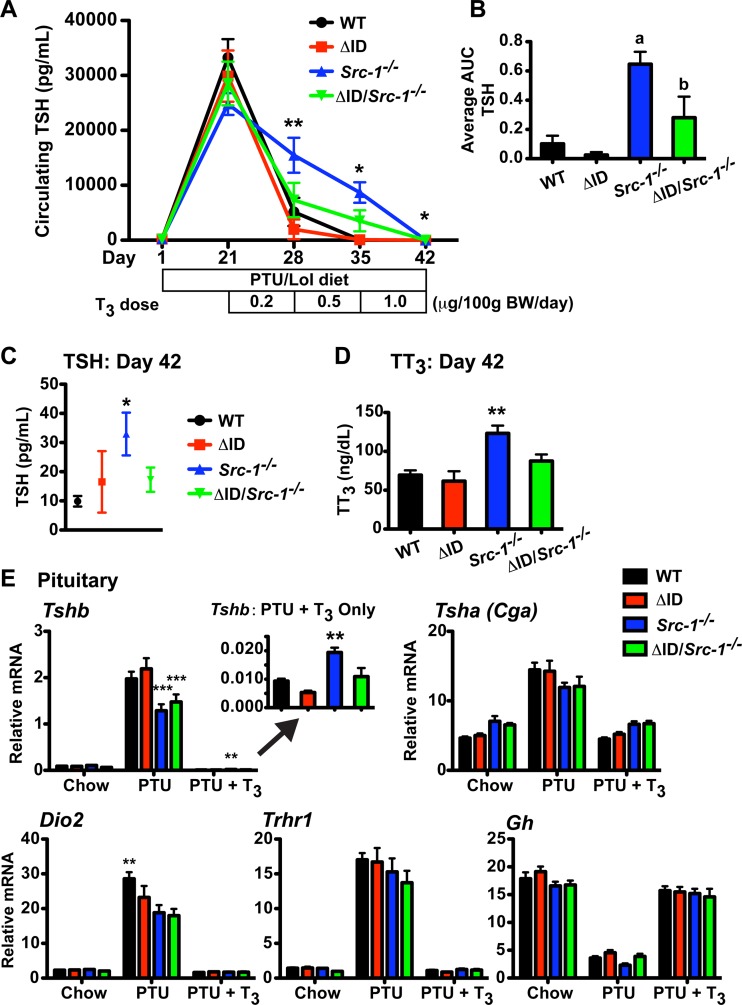
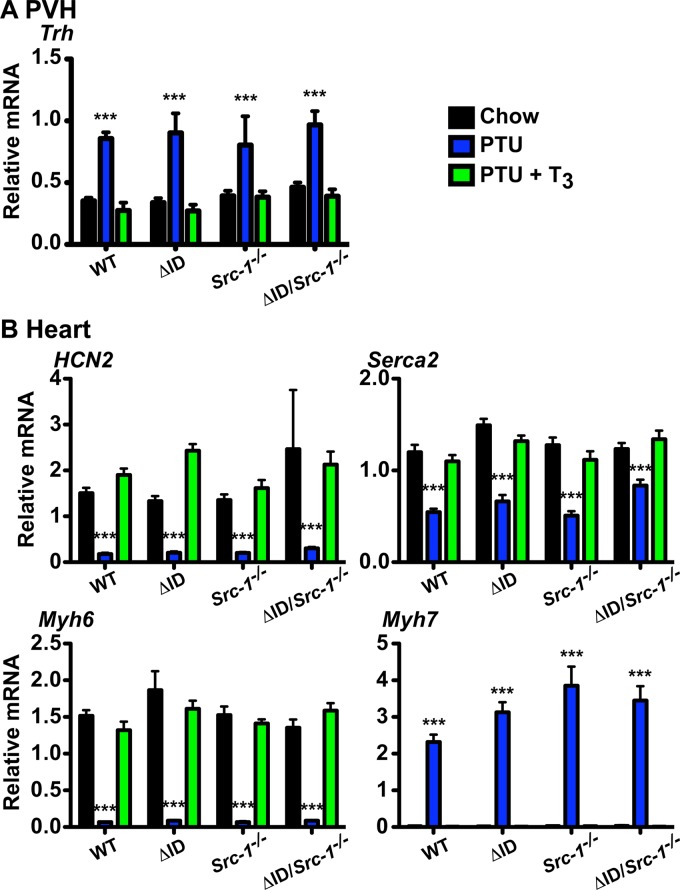
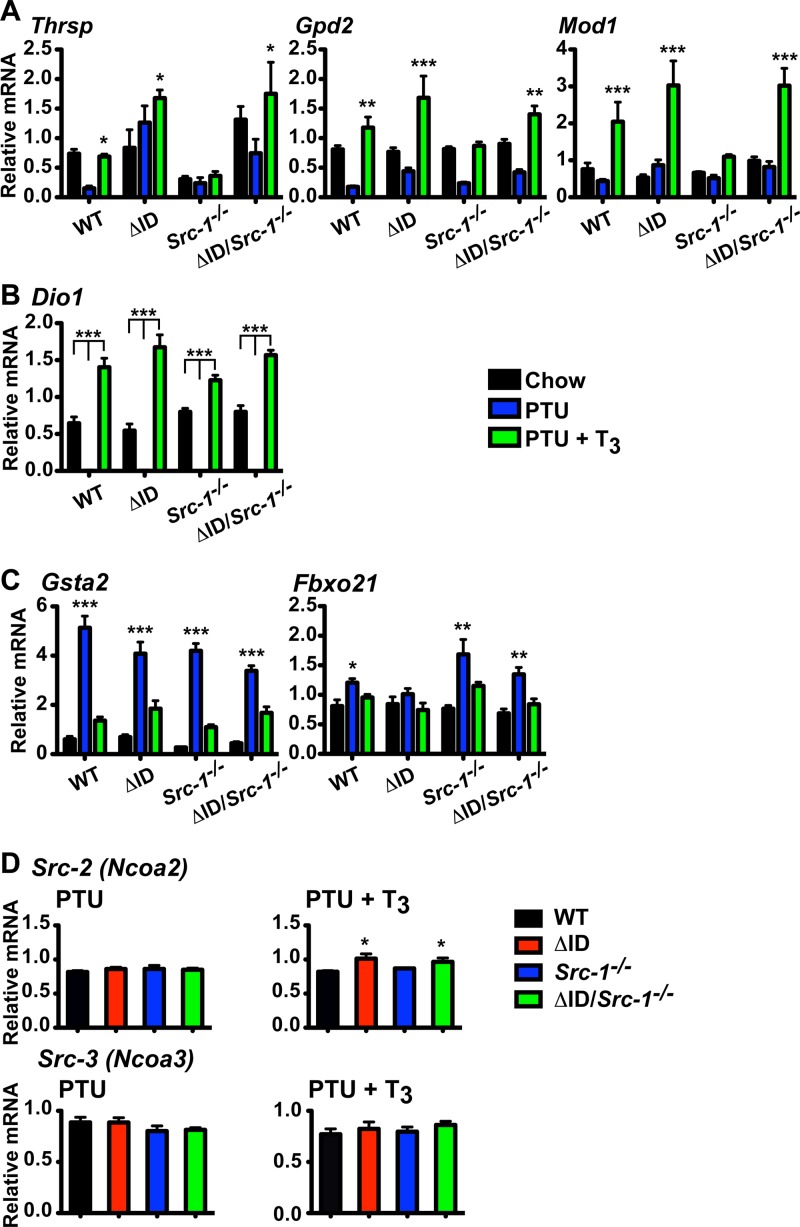
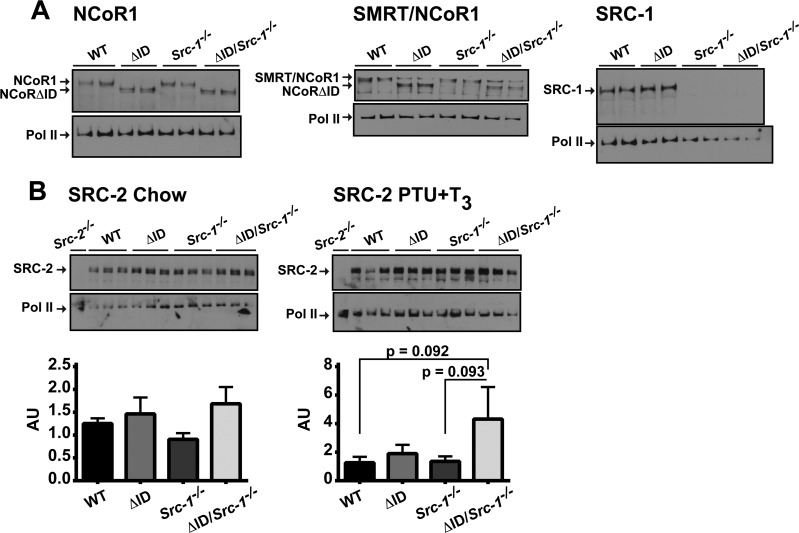
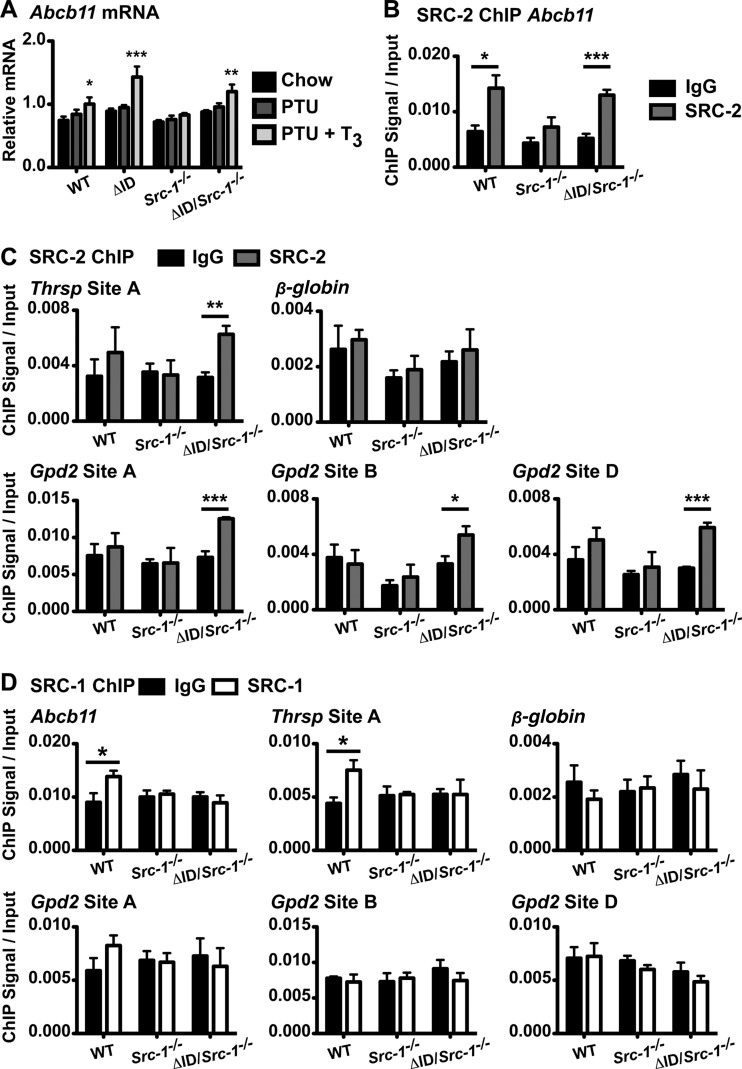
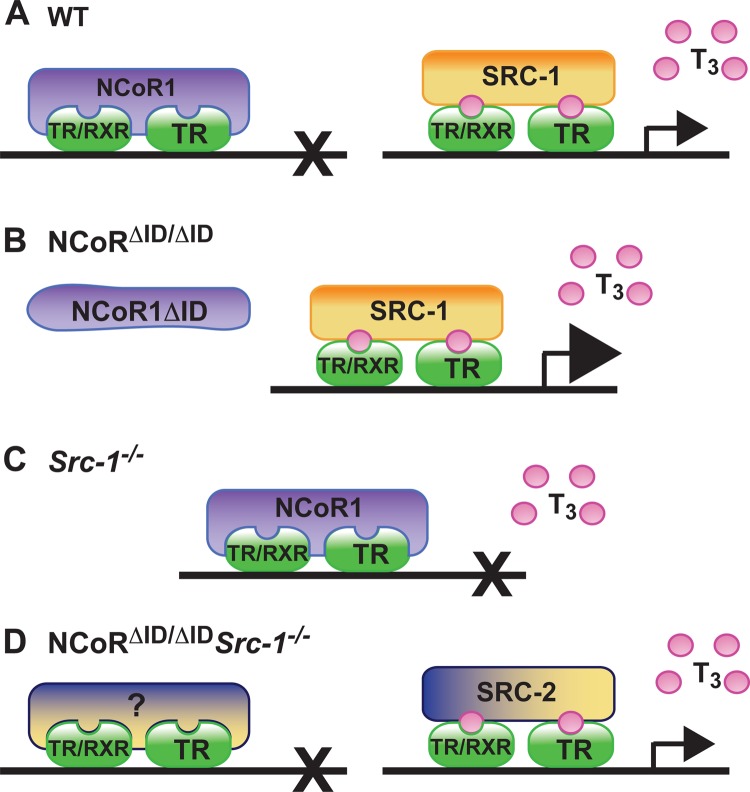
Resistance to thyroid hormone (RTH), a human syndrome, is characterized by high thyroid hormone (TH) and thyroid-stimulating hormone (TSH) levels. Mice with mutations in the thyroid hormone receptor beta (TRβ) gene that cannot bind steroid receptor coactivator 1 (SRC-1) and Src-1(-/-) mice both have phenotypes similar to that of RTH. Conversely, mice expressing a mutant nuclear corepressor 1 (Ncor1) allele that cannot interact with TRβ, termed NCoRΔID, have low TH levels and normal TSH. We hypothesized that Src-1(-/-) mice have RTH due to unopposed corepressor action. To test this, we crossed NCoRΔID and Src-1(-/-) mice to create mice deficient for coregulator action in all cell types. Remarkably, NCoR(ΔID/ΔID) Src-1(-/-) mice have normal TH and TSH levels and are triiodothryonine (T(3)) sensitive at the level of the pituitary. Although absence of SRC-1 prevented T(3) activation of key hepatic gene targets, NCoR(ΔID/ΔID) Src-1(-/-) mice reacquired hepatic T(3) sensitivity. Using in vivo chromatin immunoprecipitation assays (ChIP) for the related coactivator SRC-2, we found enhanced SRC-2 recruitment to TR-binding regions of genes in NCoR(ΔID/ΔID) Src-1(-/-) mice, suggesting that SRC-2 is responsible for T(3) sensitivity in the absence of NCoR1 and SRC-1. Thus, T(3) targets require a critical balance between NCoR1 and SRC-1. Furthermore, replacement of NCoR1 with NCoRΔID corrects RTH in Src-1(-/-) mice through increased SRC-2 recruitment to T(3) target genes.
Figures
References
-
- Yen PM. 2001. Physiological and molecular basis of thyroid hormone action. Physiol. Rev. 81:1097–1142 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous