

Molecular identification of venous progenitors in the dorsal aorta reveals an aortic origin for the cardinal vein in mammals
- PMID: 24550118
- PMCID: PMC3929407
- DOI: 10.1242/dev.101808
Molecular identification of venous progenitors in the dorsal aorta reveals an aortic origin for the cardinal vein in mammals
Abstract
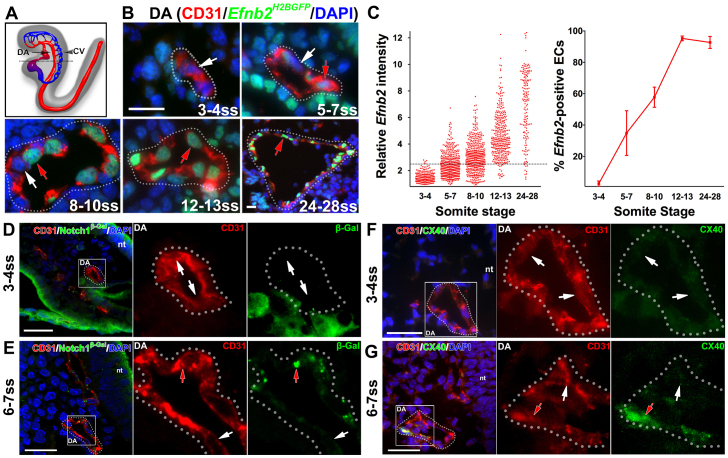
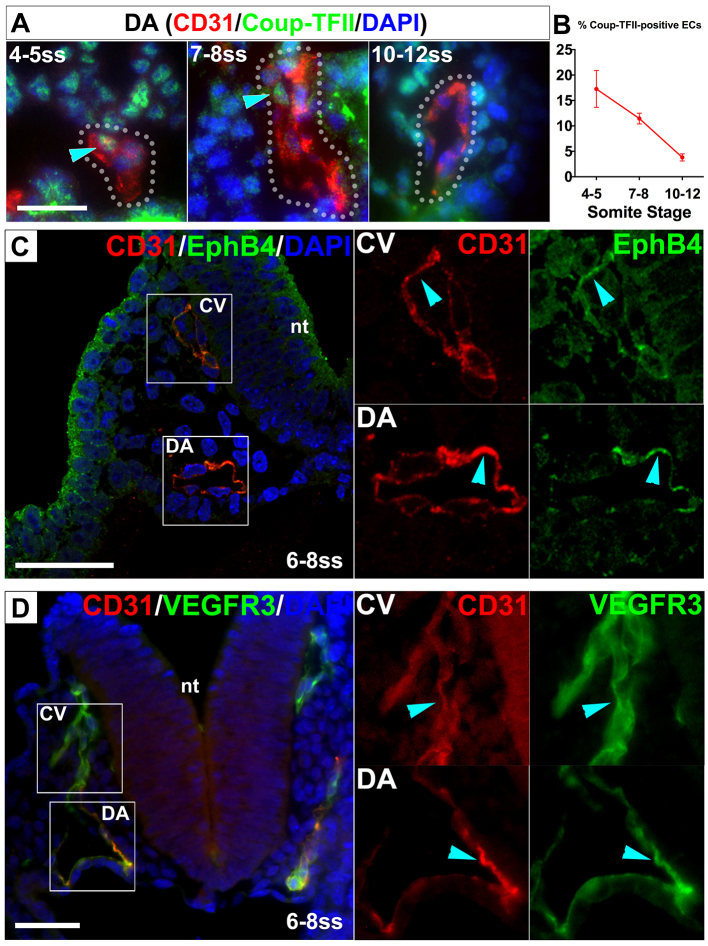
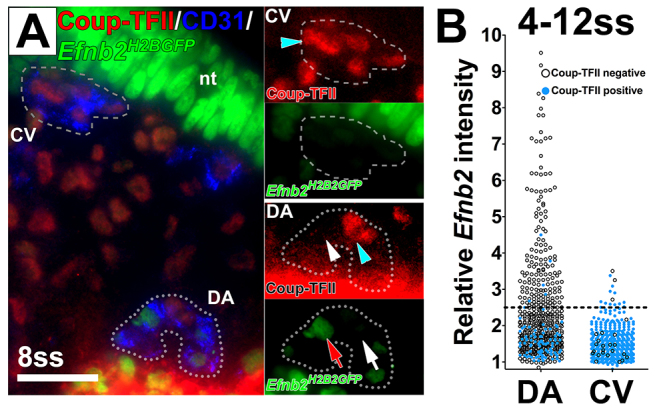
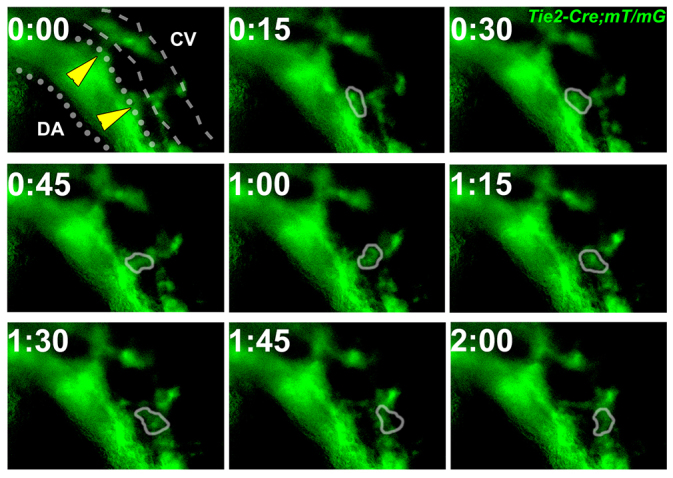
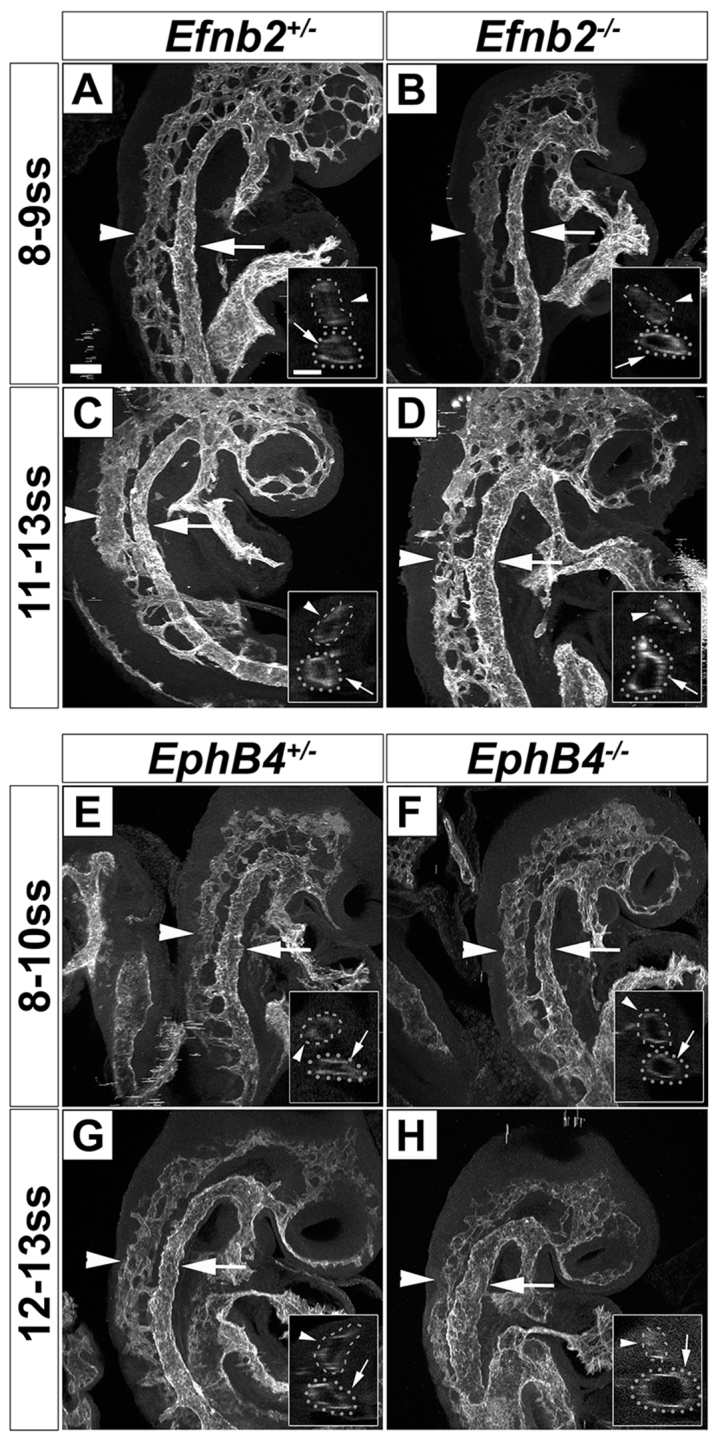
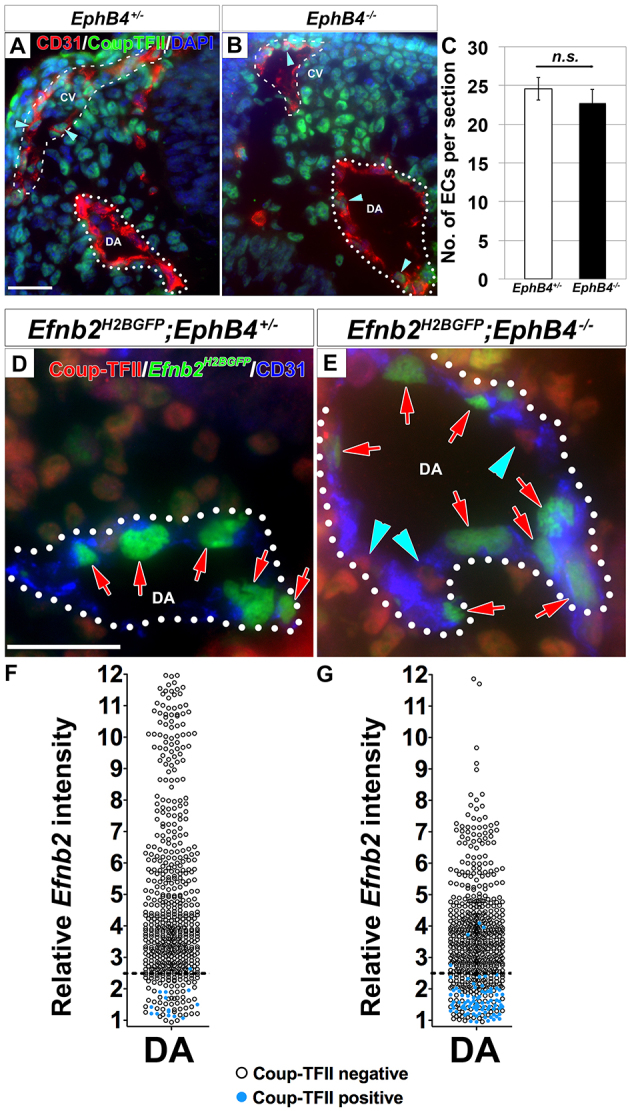
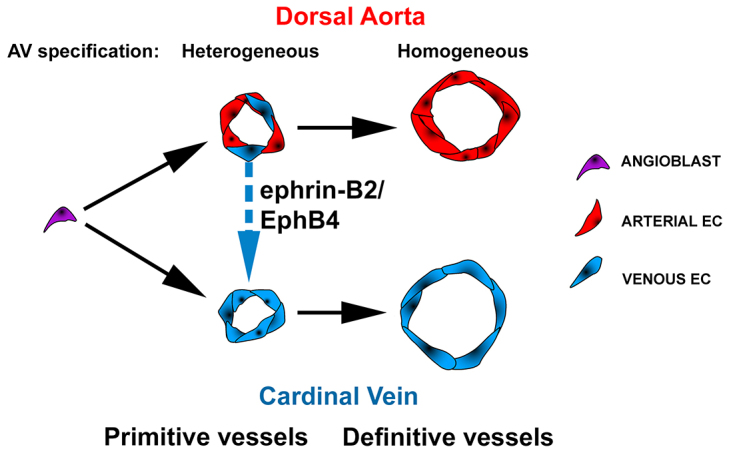
Coordinated arterial-venous differentiation is crucial for vascular development and function. The origin of the cardinal vein (CV) in mammals is unknown, while conflicting theories have been reported in chick and zebrafish. Here, we provide the first molecular characterization of endothelial cells (ECs) expressing venous molecular markers, or venous-fated ECs, within the emergent dorsal aorta (DA). These ECs, expressing the venous molecular markers Coup-TFII and EphB4, cohabited the early DA with ECs expressing the arterial molecular markers ephrin B2, Notch and connexin 40. These mixed ECs in the early DA expressed either the arterial or venous molecular marker, but rarely both. Subsequently, the DA exhibited uniform arterial markers. Real-time imaging of mouse embryos revealed EC movement from the DA to the CV during the stage when venous-fated ECs occupied the DA. We analyzed mutants for EphB4, which encodes a receptor tyrosine kinase for the ephrin B2 ligand, as we hypothesized that ephrin B2/EphB4 signaling may mediate the repulsion of venous-fated ECs from the DA to the CV. Using an EC quantification approach, we discovered that venous-fated ECs increased in the DA and decreased in the CV in the mutants, whereas the rest of the ECs in each vessel were unaffected. This result suggests that the venous-fated ECs were retained in the DA and missing in the CV in the EphB4 mutant, and thus that ephrin B2/EphB4 signaling normally functions to clear venous-fated ECs from the DA to the CV by cell repulsion. Therefore, our cellular and molecular evidence suggests that the DA harbors venous progenitors that move to participate in CV formation, and that ephrin B2/EphB4 signaling regulates this aortic contribution to the mammalian CV.
Keywords: Angiogenesis; Arterial-venous differentiation; Coup-TFII; EphB4; Ephrin B2; Mouse; Notch; Vascular development.
Figures
References
-
- Adams R. H., Wilkinson G. A., Weiss C., Diella F., Gale N. W., Deutsch U., Risau W., Klein R. (1999). Roles of ephrinB ligands and EphB receptors in cardiovascular development: demarcation of arterial/venous domains, vascular morphogenesis, and sprouting angiogenesis. Genes Dev. 13, 295–306 - PMC - PubMed
-
- Carpenter B., Lin Y., Stoll S., Raffai R. L., McCuskey R., Wang R. (2005). VEGF is crucial for the hepatic vascular development required for lipoprotein uptake. Development 132, 3293–3303 - PubMed
-
- Coffin J. D., Poole T. J. (1988). Embryonic vascular development: immunohistochemical identification of the origin and subsequent morphogenesis of the major vessel primordia in quail embryos. Development 102, 735–748 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
