

Back to front: cerebellar connections and interactions with the prefrontal cortex
- PMID: 24550789
- PMCID: PMC3912388
- DOI: 10.3389/fnsys.2014.00004
Back to front: cerebellar connections and interactions with the prefrontal cortex
Abstract
Although recent neuroanatomical evidence has demonstrated closed-loop connectivity between prefrontal cortex and the cerebellum, the physiology of cerebello-cerebral circuits and the extent to which cerebellar output modulates neuronal activity in neocortex during behavior remain relatively unexplored. We show that electrical stimulation of the contralateral cerebellar fastigial nucleus (FN) in awake, behaving rats evokes distinct local field potential (LFP) responses (onset latency ~13 ms) in the prelimbic (PrL) subdivision of the medial prefrontal cortex. Trains of FN stimulation evoke heterogeneous patterns of response in putative pyramidal cells in frontal and prefrontal regions in both urethane-anesthetized and awake, behaving rats. However, the majority of cells showed decreased firing rates during stimulation and subsequent rebound increases; more than 90% of cells showed significant changes in response. Simultaneous recording of on-going LFP activity from FN and PrL while rats were at rest or actively exploring an open field arena revealed significant network coherence restricted to the theta frequency range (5-10 Hz). Granger causality analysis indicated that this coherence was significantly directed from cerebellum to PrL during active locomotion. Our results demonstrate the presence of a cerebello-prefrontal pathway in rat and reveal behaviorally dependent coordinated network activity between the two structures, which could facilitate transfer of sensorimotor information into ongoing neocortical processing during goal directed behaviors.
Keywords: cerebellum; coherence; fastigial nucleus; prefrontal cortex; prelimbic cortex; theta.
Figures
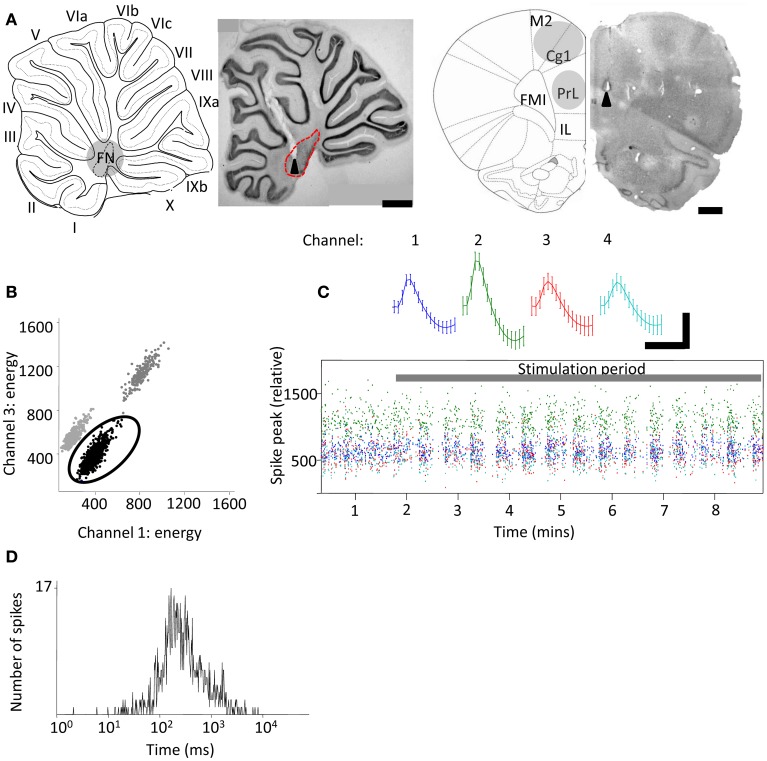

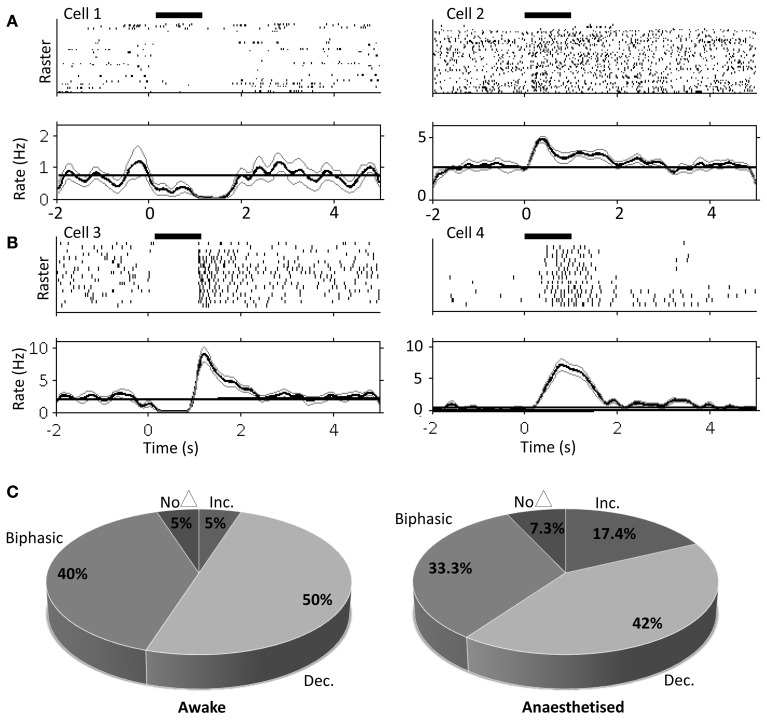

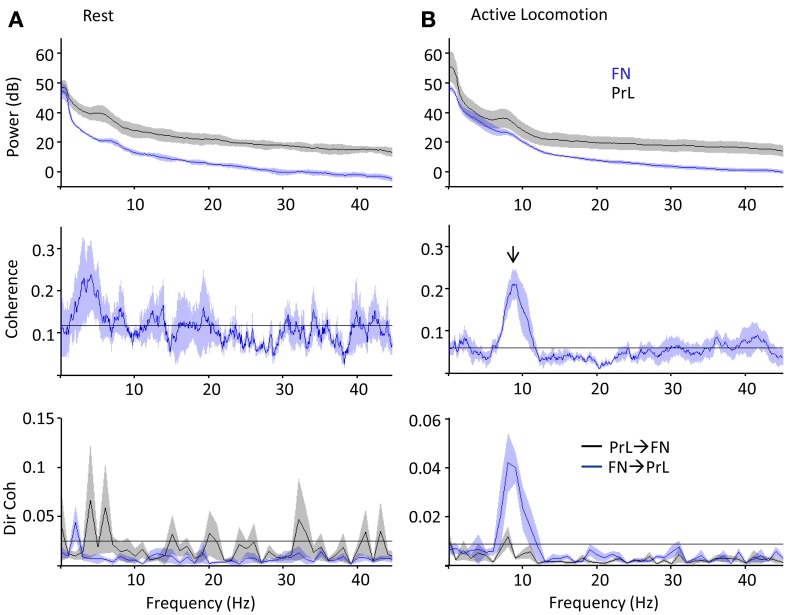
References
-
- Andreasen N. C., O'Leary D. S., Cizadlo T., Arndt S., Rezai K., Ponto L. L., et al. (1996). Schizophrenia and cognitive dysmetria: a positron-emission tomography study of dysfunctional prefrontal-thalamic-cerebellar circuitry. Proc. Natl. Acad. Sci. U.S.A. 93, 9985–9990 10.1073/pnas.93.18.9985 - DOI - PMC - PubMed
-
- Arguello P. A., Enquist L. W., Wang S. S.-H. (2012). Long-distance connectivity between prefrontal cortex and cerebellum in mouse, in Society for Neuroscience (New Orleans: ).
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous