

The emerging roles of ribosome biogenesis in craniofacial development
- PMID: 24550838
- PMCID: PMC3912750
- DOI: 10.3389/fphys.2014.00026
The emerging roles of ribosome biogenesis in craniofacial development
Abstract
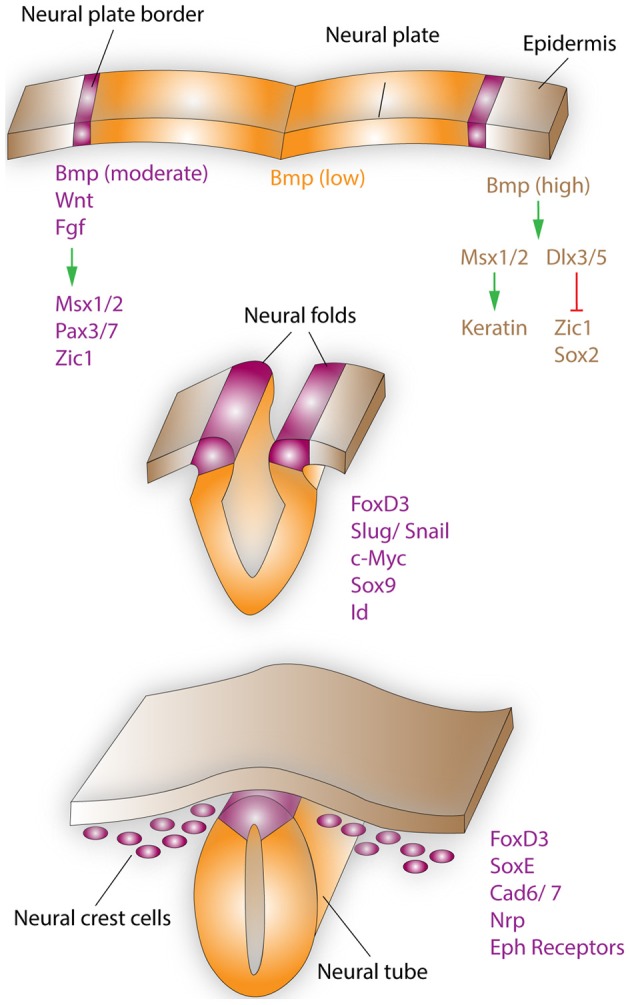
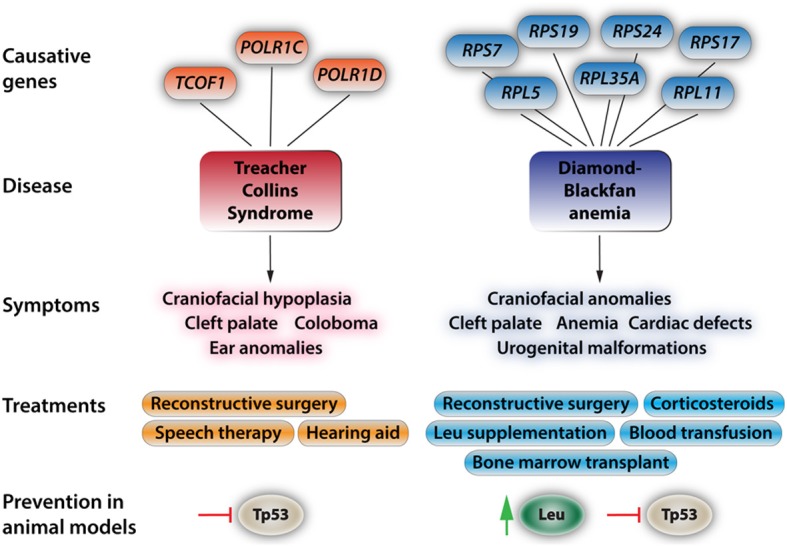
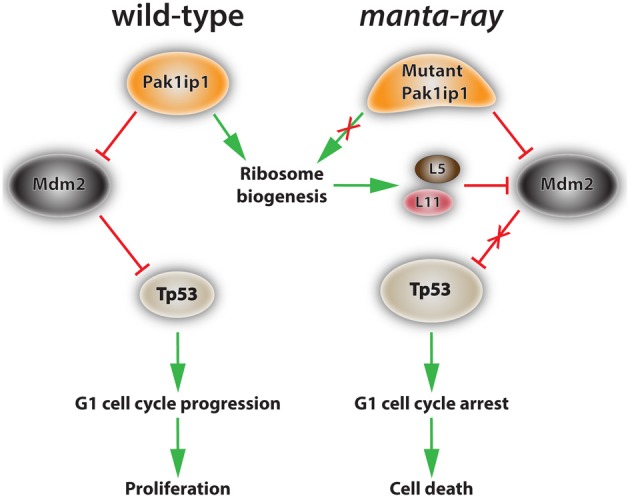
Neural crest cells (NCCs) are a transient, migratory cell population, which originates during neurulation at the neural folds and contributes to the majority of tissues, including the mesenchymal structures of the craniofacial skeleton. The deregulation of the complex developmental processes that guide migration, proliferation, and differentiation of NCCs may result in a wide range of pathological conditions grouped together as neurocristopathies. Recently, due to their multipotent properties neural crest stem cells have received considerable attention as a possible source for stem cell based regenerative therapies. This exciting prospect underlines the need to further explore the developmental programs that guide NCC differentiation. This review explores the particular importance of ribosome biogenesis defects in this context since a specific interface between ribosomopathies and neurocristopathies exists as evidenced by disorders such as Treacher-Collins-Franceschetti syndrome (TCS) and Diamond-Blackfan anemia (DBA).
Keywords: TP53; cell cycle regulation; craniofacial development; neural crest; neurocristopathies; ribosome biogenesis; ribosomopathies.
Figures
References
-
- Abu-Issa R., Smyth G., Smoak I., Yamamura K., Meyers E. N. (2002). Fgf8 is required for pharyngeal arch and cardiovascular development in the mouse. Development 129, 4613–4625 - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous