

The NAD+ synthesizing enzyme nicotinamide mononucleotide adenylyltransferase 2 (NMNAT-2) is a p53 downstream target
- PMID: 24552824
- PMCID: PMC3984302
- DOI: 10.4161/cc.28128
The NAD+ synthesizing enzyme nicotinamide mononucleotide adenylyltransferase 2 (NMNAT-2) is a p53 downstream target
Abstract
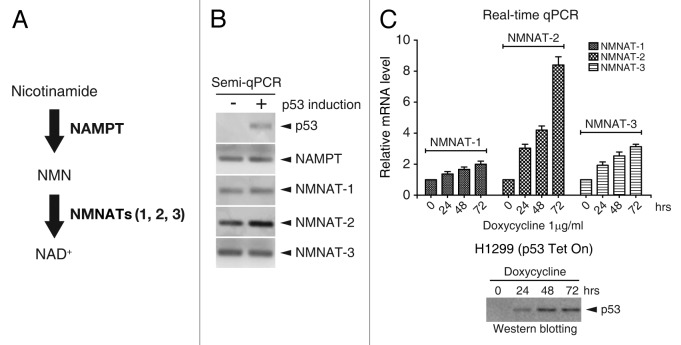
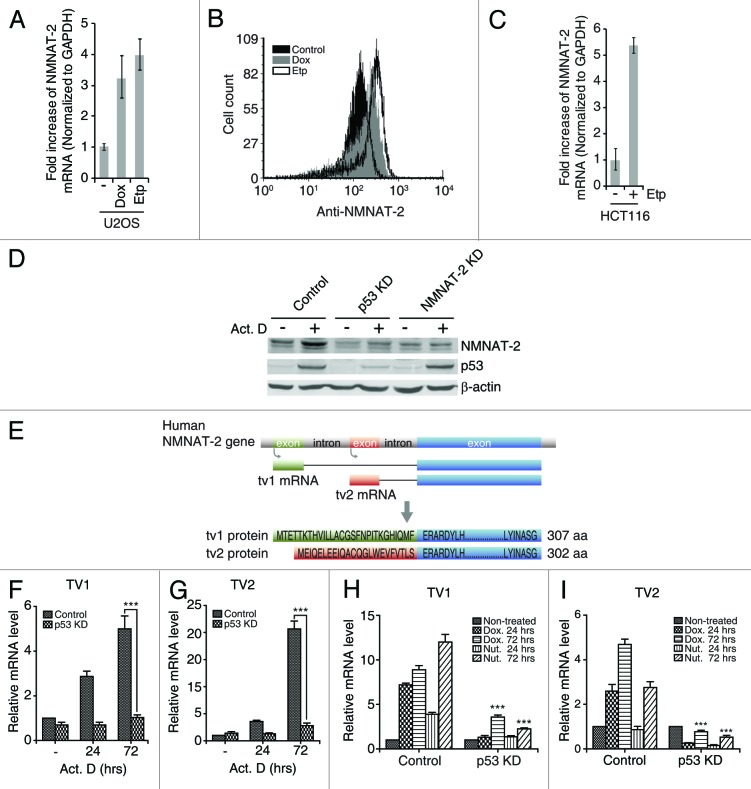
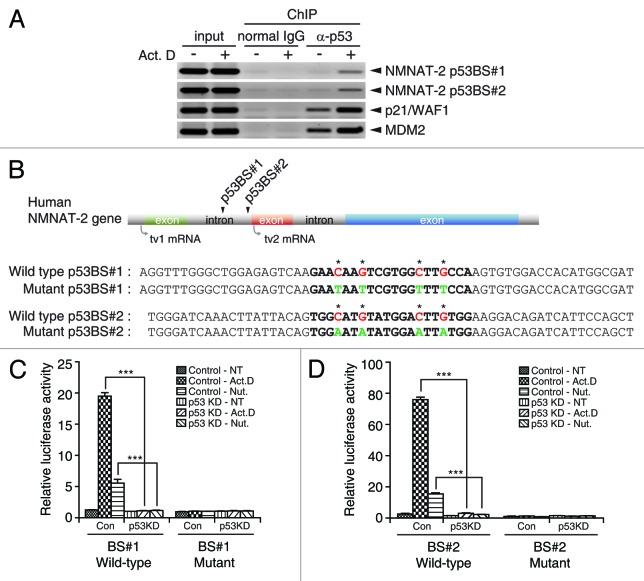
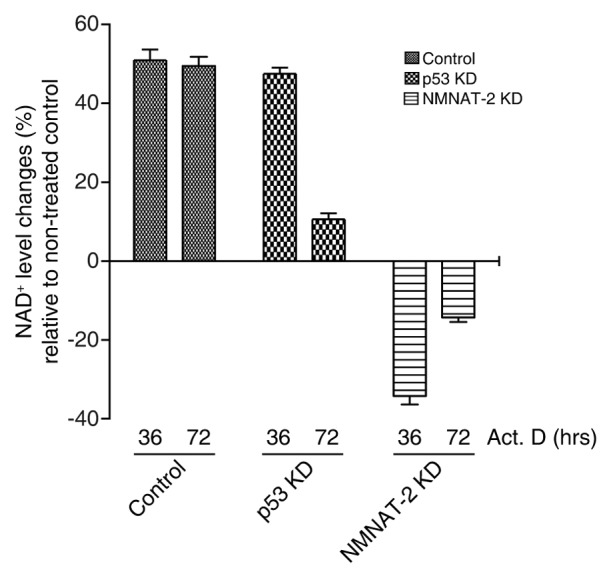
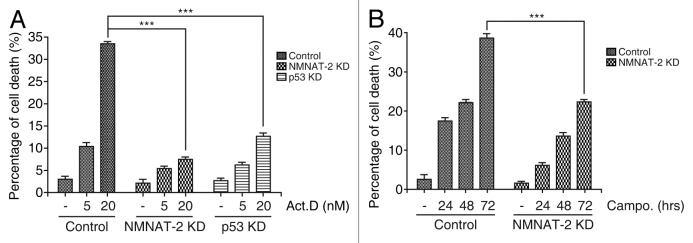
NAD(+) metabolism plays key roles not only in energy production but also in diverse cellular physiology. Aberrant NAD(+) metabolism is considered a hallmark of cancer. Recently, the tumor suppressor p53, a major player in cancer signaling pathways, has been implicated as an important regulator of cellular metabolism. This notion led us to examine whether p53 can regulate NAD(+) biosynthesis in the cell. Our search resulted in the identification of nicotinamide mononucleotide adenylyltransferase 2 (NMNAT-2), a NAD(+) synthetase, as a novel downstream target gene of p53. We show that NMNAT-2 expression is induced upon DNA damage in a p53-dependent manner. Two putative p53 binding sites were identified within the human NMNAT-2 gene, and both were found to be functional in a p53-dependent manner. Furthermore, knockdown of NMNAT-2 significantly reduces cellular NAD(+) levels and protects cells from p53-dependent cell death upon DNA damage, suggesting an important functional role of NMNAT-2 in p53-mediated signaling. Our demonstration that p53 modulates cellular NAD(+) synthesis is congruent with p53's emerging role as a key regulator of metabolism and related cell fate.
Keywords: NAD+ biosynthesis; NMNAT-2; apoptosis; p53.
Figures
Comment in
-
p53, DNA damage, and NAD+ homeostasis.Cell Cycle. 2014;13(11):1661-2. doi: 10.4161/cc.29151. Epub 2014 May 8. Cell Cycle. 2014. PMID: 24810733 Free PMC article. No abstract available.
References
-
- Di Stefano G, Manerba M, Vettraino M. NAD metabolism and functions: a common therapeutic target for neoplastic, metabolic and neurodegenerative diseases. Curr Top Med Chem. 2013;13:2918–29. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous