

Caenorhabditis elegans oocyte meiotic spindle pole assembly requires microtubule severing and the calponin homology domain protein ASPM-1
- PMID: 24554763
- PMCID: PMC3982995
- DOI: 10.1091/mbc.E13-11-0687
Caenorhabditis elegans oocyte meiotic spindle pole assembly requires microtubule severing and the calponin homology domain protein ASPM-1
Abstract
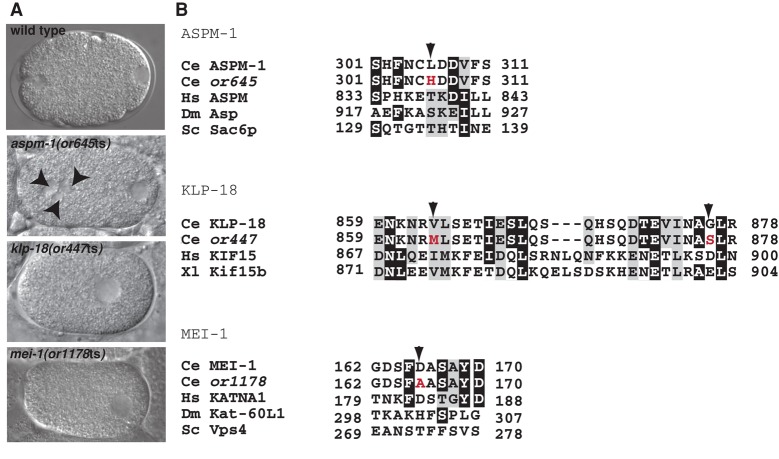
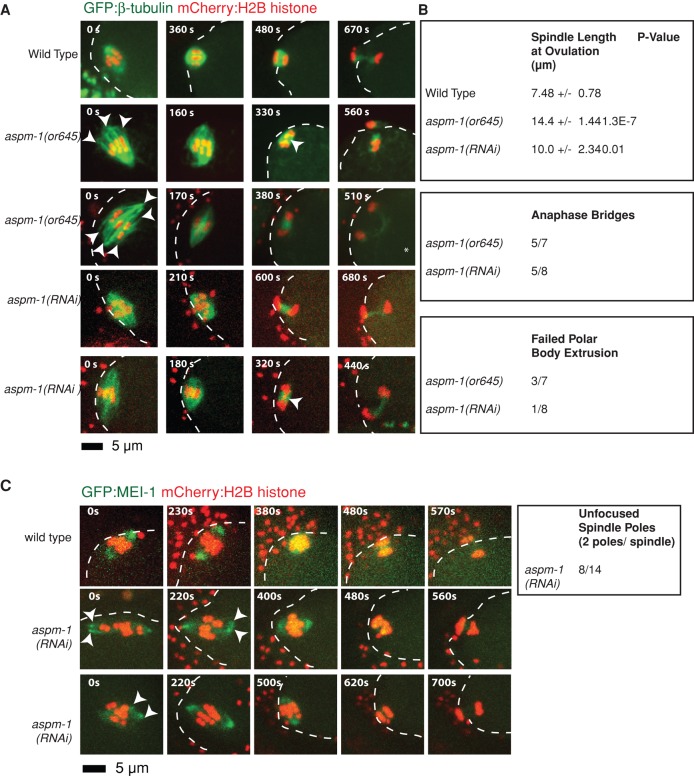
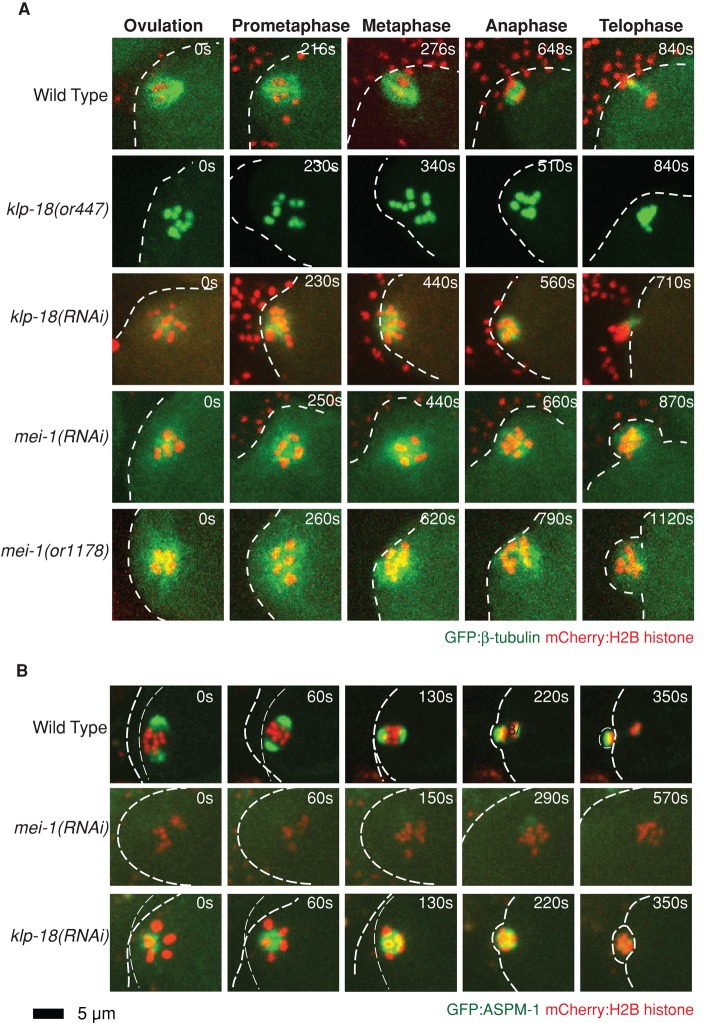
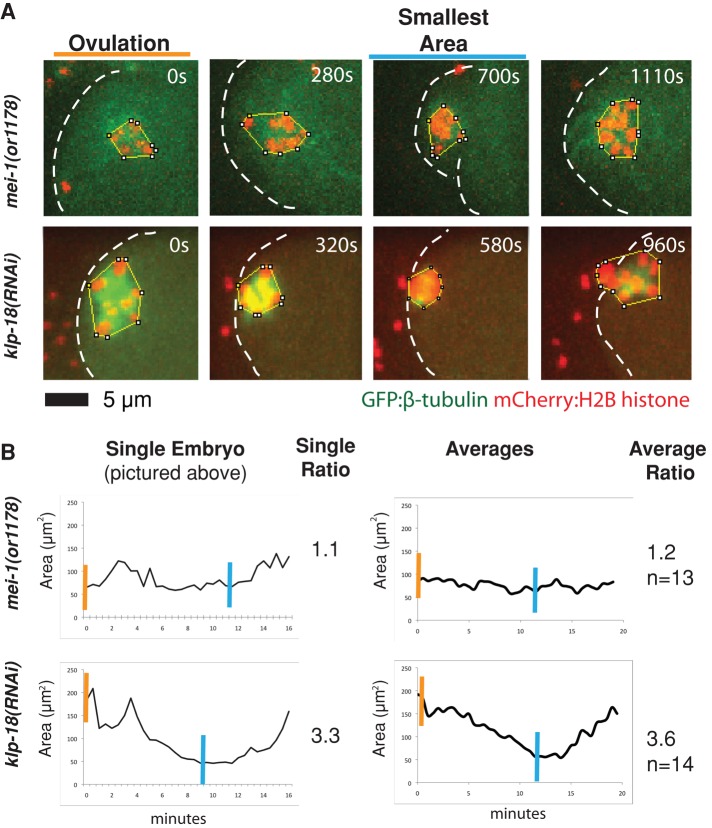
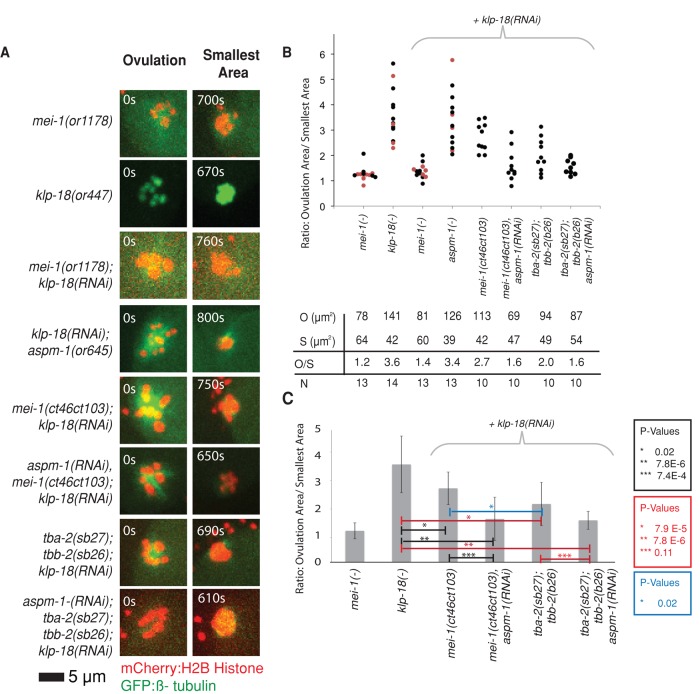
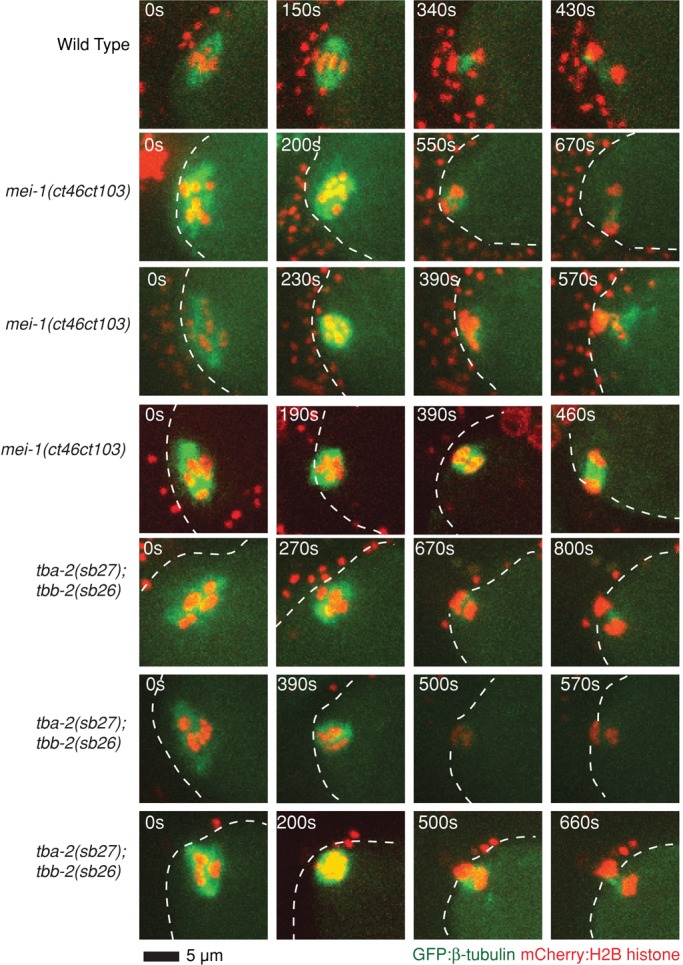
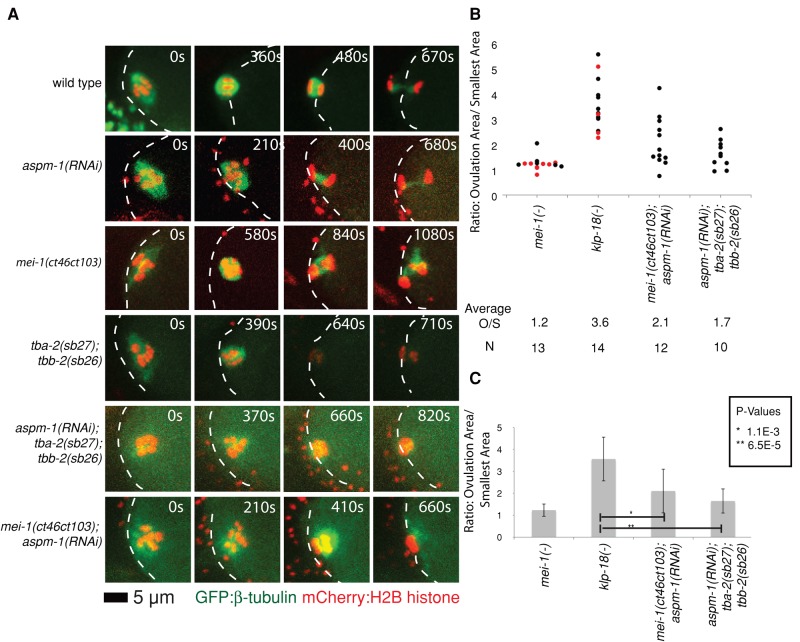
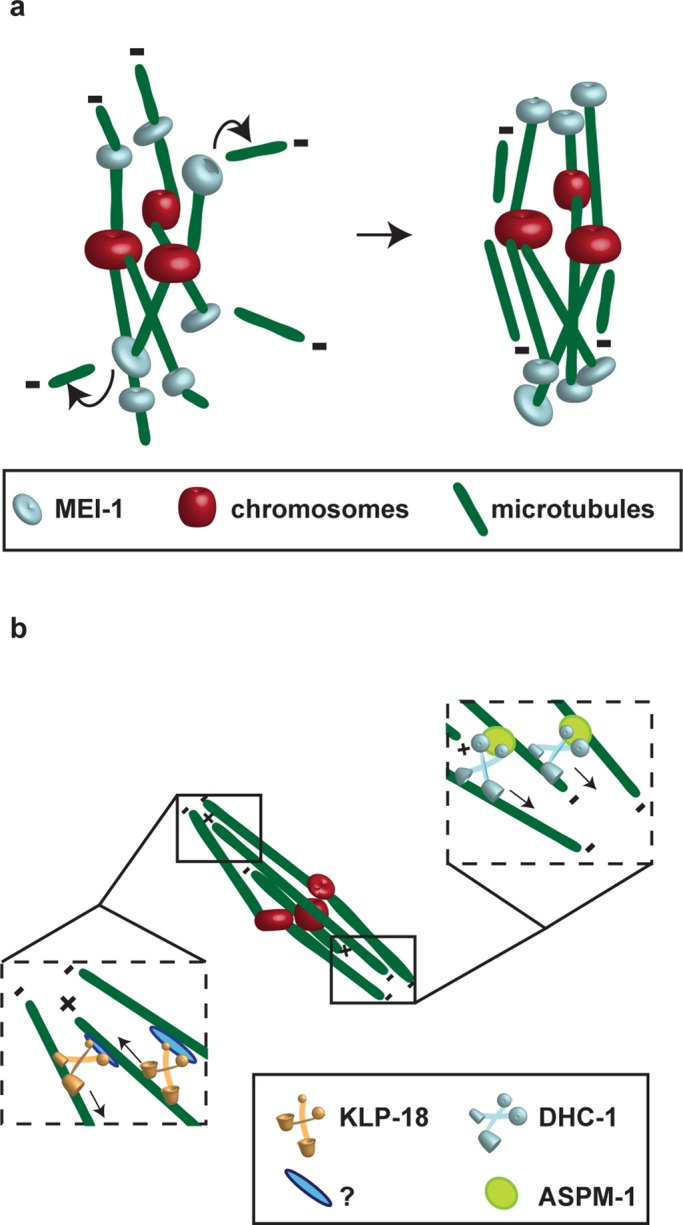
In many animals, including vertebrates, oocyte meiotic spindles are bipolar but assemble in the absence of centrosomes. Although meiotic spindle positioning in oocytes has been investigated extensively, much less is known about their assembly. In Caenorhabditis elegans, three genes previously shown to contribute to oocyte meiotic spindle assembly are the calponin homology domain protein encoded by aspm-1, the katanin family member mei-1, and the kinesin-12 family member klp-18. We isolated temperature-sensitive alleles of all three and investigated their requirements using live-cell imaging to reveal previously undocumented requirements for aspm-1 and mei-1. Our results indicate that bipolar but abnormal oocyte meiotic spindles assemble in aspm-1(-) embryos, whereas klp-18(-) and mei-1(-) mutants assemble monopolar and apolar spindles, respectively. Furthermore, two MEI-1 functions--ASPM-1 recruitment to the spindle and microtubule severing--both contribute to monopolar spindle assembly in klp-18(-) mutants. We conclude that microtubule severing and ASPM-1 both promote meiotic spindle pole assembly in C. elegans oocytes, whereas the kinesin 12 family member KLP-18 promotes spindle bipolarity.
Figures
References
-
- Albertson DG. Formation of the first cleavage spindle in nematode embryos. Dev Biol. 1984;101:61–72. - PubMed
-
- Albertson DG, Thomson JN. Segregation of holocentric chromosomes at meiosis in the nematode, Caenorhabditis elegans. Chromosome Res. 1993;1:15–26. - PubMed
-
- Bond J, et al. ASPM is a major determinant of cerebral cortical size. Nat Genet. 2002;32:316–320. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
