

Upregulation of capacity for glutathione synthesis in response to amino acid deprivation: regulation of glutamate-cysteine ligase subunits
- PMID: 24557597
- PMCID: PMC3991118
- DOI: 10.1007/s00726-014-1687-1
Upregulation of capacity for glutathione synthesis in response to amino acid deprivation: regulation of glutamate-cysteine ligase subunits
Abstract
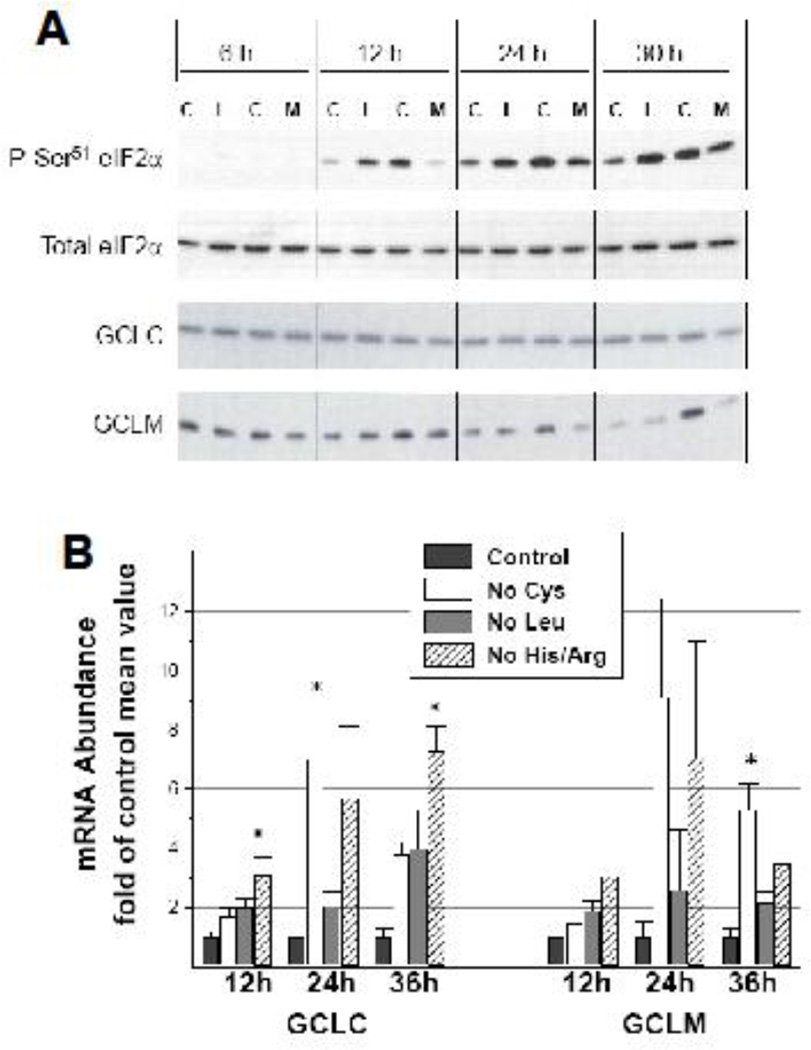
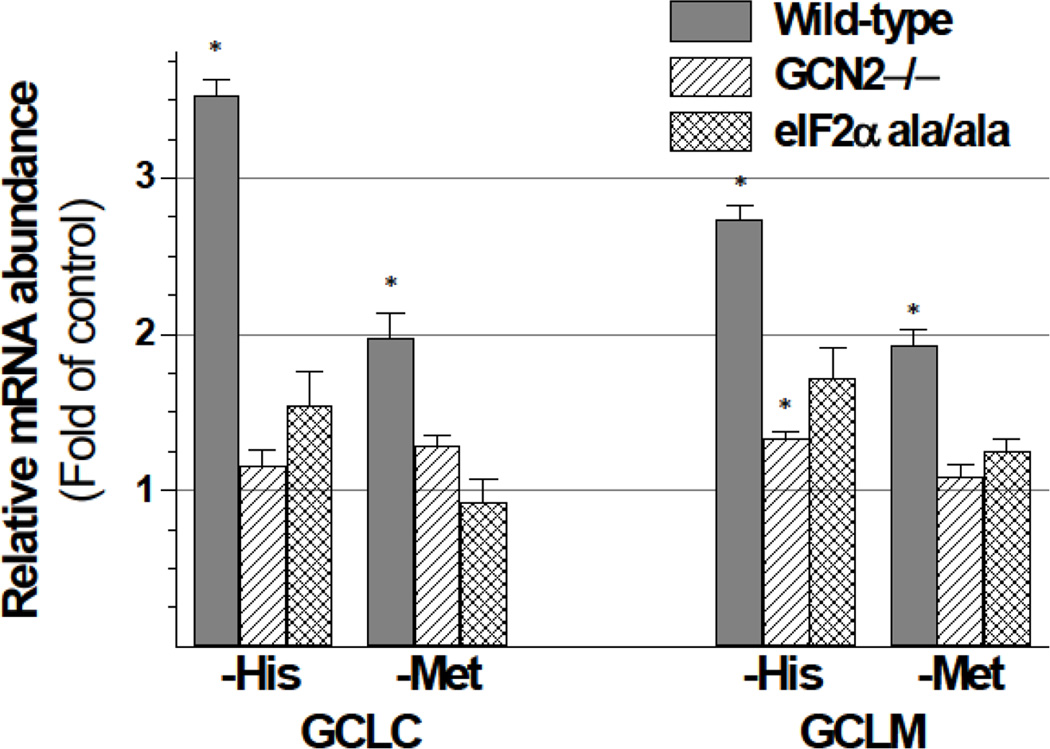
Using HepG2/C3A cells and MEFs, we investigated whether induction of GSH synthesis in response to sulfur amino acid deficiency is mediated by the decrease in cysteine levels or whether it requires a decrease in GSH levels per se. Both the glutamate-cysteine ligase catalytic (GCLC) and modifier (GCLM) subunit mRNA levels were upregulated in response to a lack of cysteine or other essential amino acids, independent of GSH levels. This upregulation did not occur in MEFs lacking GCN2 (general control non-derepressible 2, also known as eIF2α kinase 4) or in cells expressing mutant eIF2α lacking the eIF2α kinase Ser(51) phosphorylation site, indicating that expression of both GCLC and GCLM was mediated by the GCN2/ATF4 stress response pathway. Only the increase in GCLM mRNA level, however, was accompanied by a parallel increase in protein expression, suggesting that the enhanced capacity for GSH synthesis depended largely on increased association of GCLC with its regulatory subunit. Upregulation of both GCLC and GLCM mRNA levels in response to cysteine deprivation was dependent on new protein synthesis, which is consistent with expression of GCLC and GCLM being mediated by proteins whose synthesis depends on activation of the GCN2/ATF4 pathway. Our data suggest that the regulation of GCLC expression may be mediated by changes in the abundance of transcriptional regulators, whereas the regulation of GCLM expression may be mediated by changes in the abundance of mRNA stabilizing or destabilizing proteins. Upregulation of GCLM levels in response to low cysteine levels may serve to protect the cell in the face of a future stress requiring GSH as an antioxidant or conjugating/detoxifying agent.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures





Similar articles
-
Differential regulation of glutamate-cysteine ligase subunit expression and increased holoenzyme formation in response to cysteine deprivation.Biochem J. 2006 Jan 1;393(Pt 1):181-90. doi: 10.1042/BJ20051111. Biochem J. 2006. PMID: 16137247 Free PMC article.
-
Glutamate cysteine ligase (GCL) transgenic and gene-targeted mice for controlling glutathione synthesis.Curr Protoc Toxicol. 2009 Feb;Chapter 6:Unit6.16. doi: 10.1002/0471140856.tx0616s39. Curr Protoc Toxicol. 2009. PMID: 23045016
-
Glutamate cysteine ligase catalysis: dependence on ATP and modifier subunit for regulation of tissue glutathione levels.J Biol Chem. 2005 Oct 7;280(40):33766-74. doi: 10.1074/jbc.M504604200. Epub 2005 Aug 4. J Biol Chem. 2005. PMID: 16081425
-
Modulating GSH synthesis using glutamate cysteine ligase transgenic and gene-targeted mice.Drug Metab Rev. 2008;40(3):465-77. doi: 10.1080/03602530802186587. Drug Metab Rev. 2008. PMID: 18642143 Review.
-
Structure, function, and post-translational regulation of the catalytic and modifier subunits of glutamate cysteine ligase.Mol Aspects Med. 2009 Feb-Apr;30(1-2):86-98. doi: 10.1016/j.mam.2008.08.009. Epub 2008 Sep 6. Mol Aspects Med. 2009. PMID: 18812186 Free PMC article. Review.
Cited by
-
1,4-dihydroxy quininib activates ferroptosis pathways in metastatic uveal melanoma and reveals a novel prognostic biomarker signature.Cell Death Discov. 2024 Feb 10;10(1):70. doi: 10.1038/s41420-023-01773-8. Cell Death Discov. 2024. PMID: 38341410 Free PMC article.
-
Glucose Availability Alters Gene and Protein Expression of Several Newly Classified and Putative Solute Carriers in Mice Cortex Cell Culture and D. melanogaster.Front Cell Dev Biol. 2020 Jul 7;8:579. doi: 10.3389/fcell.2020.00579. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 32733888 Free PMC article.
-
Curcumin Modulation of the Gut-Brain Axis for Neuroinflammation and Metabolic Disorders Prevention and Treatment.Nutrients. 2025 Apr 24;17(9):1430. doi: 10.3390/nu17091430. Nutrients. 2025. PMID: 40362738 Free PMC article. Review.
-
Glutathione-mediated antioxidant response and aerobic metabolism: two crucial factors involved in determining the multi-drug resistance of high-risk neuroblastoma.Oncotarget. 2016 Oct 25;7(43):70715-70737. doi: 10.18632/oncotarget.12209. Oncotarget. 2016. PMID: 27683112 Free PMC article.
-
The Minimum Methionine Requirement for Adults Aged ≥60 Years Is the Same in Males and Females.Nutrients. 2023 Sep 23;15(19):4112. doi: 10.3390/nu15194112. Nutrients. 2023. PMID: 37836396 Free PMC article.
References
-
- Anderson ME. Glutathione: an overview of biosynthesis and modulation. Chem Biol Interact. 1998;111:1–14. - PubMed
-
- Cereser C, Guichard J, Drai J, Bannier E, Garcia I, et al. Quantitation of reduced and total glutathione at the femtomole level by high-performance liquid chromatography with fluorescence detection: application to red blood cells and cultured fibroblasts. J Chromatography. 2001;752:123–132. - PubMed
-
- Chan JY, Kwong M. Impaired expression of glutathione synthetic enzyme genes in mice with targeted deletion of the Nrf2 basic-leucine zipper protein. Biochim Biophys Acta. 2000;1517:19–26. - PubMed
-
- Chen Y, Shertzer HG, Schneider SN, Nebert DW, Dalton TP. Glutamate cysteine ligase catalysis: dependence on ATP and modifier subunit for regulation of tissue glutathione levels. J Biol Chem. 2005;280:33766–33774. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
