

The perforin pore facilitates the delivery of cationic cargos
- PMID: 24558045
- PMCID: PMC3979413
- DOI: 10.1074/jbc.M113.544890
The perforin pore facilitates the delivery of cationic cargos
Abstract
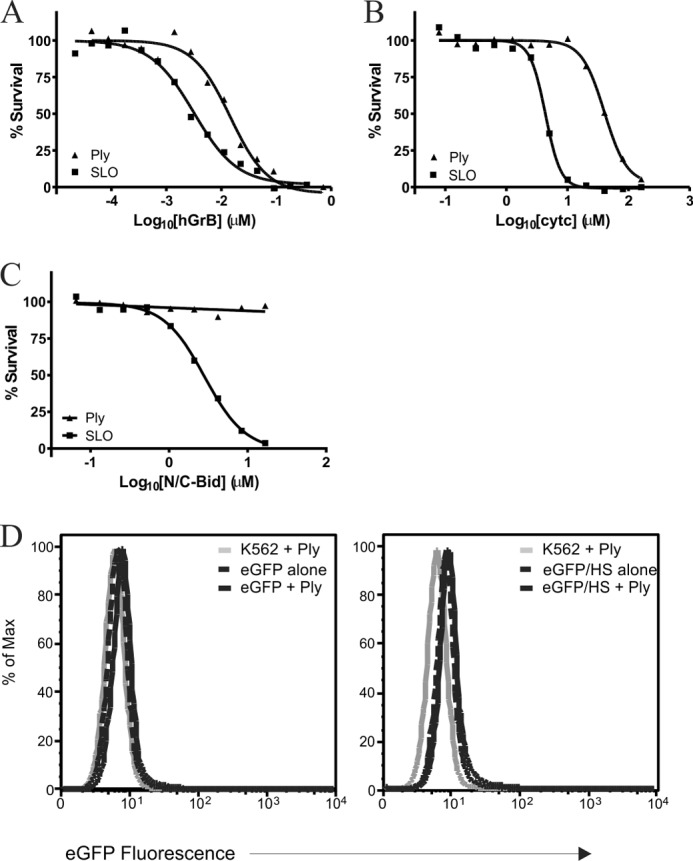
Cytotoxic lymphocytes eliminate virally infected or neoplastic cells through the action of cytotoxic proteases (granzymes). The pore-forming protein perforin is essential for delivery of granzymes into the cytoplasm of target cells; however the mechanism of this delivery is incompletely understood. Perforin contains a membrane attack complex/perforin (MACPF) domain and oligomerizes to form an aqueous pore in the plasma membrane; therefore the simplest (and best supported) model suggests that granzymes passively diffuse through the perforin pore into the cytoplasm of the target cell. Here we demonstrate that perforin preferentially delivers cationic molecules while anionic and neutral cargoes are delivered inefficiently. Furthermore, another distantly related pore-forming MACPF protein, pleurotolysin (from the oyster mushroom), also favors the delivery of cationic molecules, and efficiently delivers human granzyme B. We propose that this facilitated diffusion is due to conserved features of oligomerized MACPF proteins, which may include an anionic lumen.
Keywords: Cell Death; Cell Permeabilization; Cellular Immune Response; Granzyme; MACPF; Natural Killer (NK) Cell; Perforin; Pore; Streptolysin O; T Cell Biology.
Figures




References
-
- Metkar S. S., Menaa C., Pardo J., Wang B., Wallich R., Freudenberg M., Kim S., Raja S. M., Shi L., Simon M. M., Froelich C. J. (2008) Human and mouse granzyme A induce a proinflammatory cytokine response. Immunity 29, 720–733 - PubMed
-
- Kägi D., Ledermann B., Bürki K., Seiler P., Odermatt B., Olsen K. J., Podack E. R., Zinkernagel R. M., Hengartner H. (1994) Cytotoxicity mediated by T cells and natural killer cells is greatly impaired in perforin-deficient mice. Nature 369, 31–37 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
