

The transcriptional complex between the BCL2 i-motif and hnRNP LL is a molecular switch for control of gene expression that can be modulated by small molecules
- PMID: 24559432
- PMCID: PMC3985447
- DOI: 10.1021/ja4109352
The transcriptional complex between the BCL2 i-motif and hnRNP LL is a molecular switch for control of gene expression that can be modulated by small molecules
Abstract
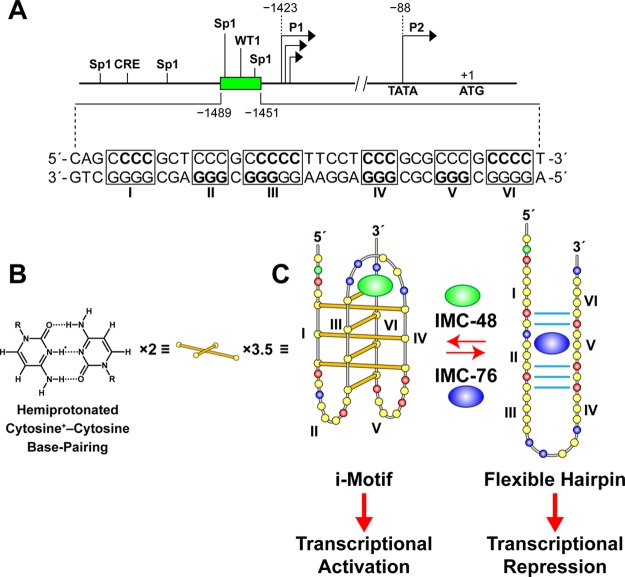
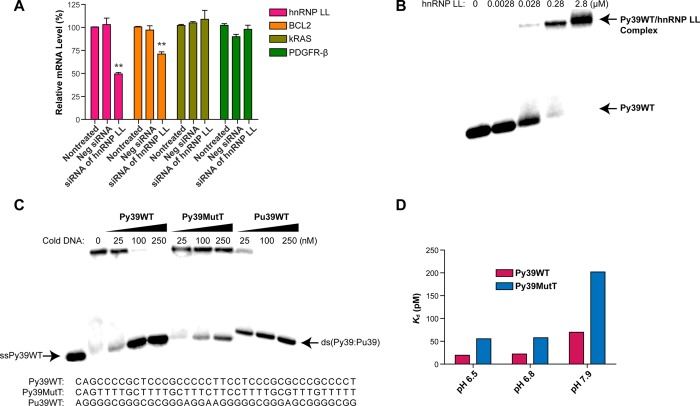
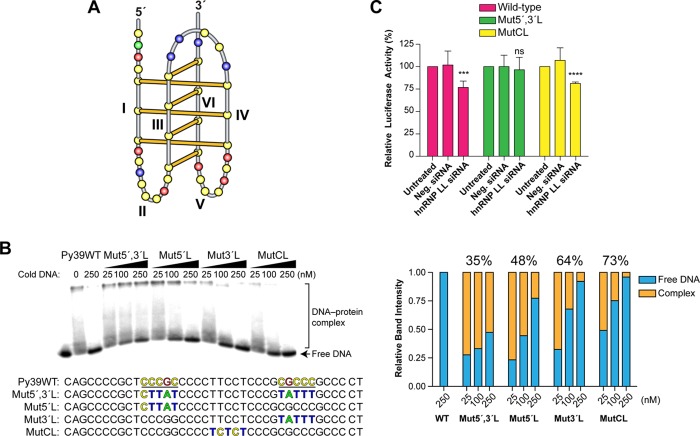
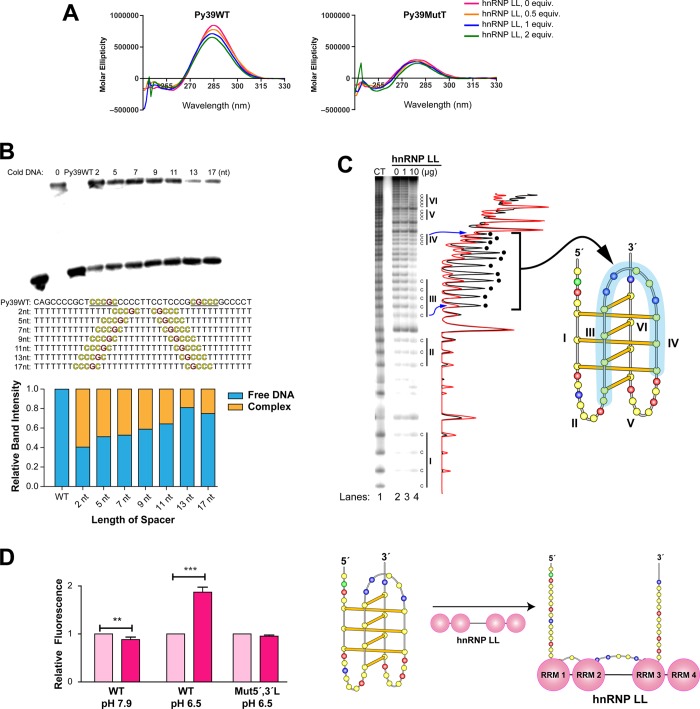
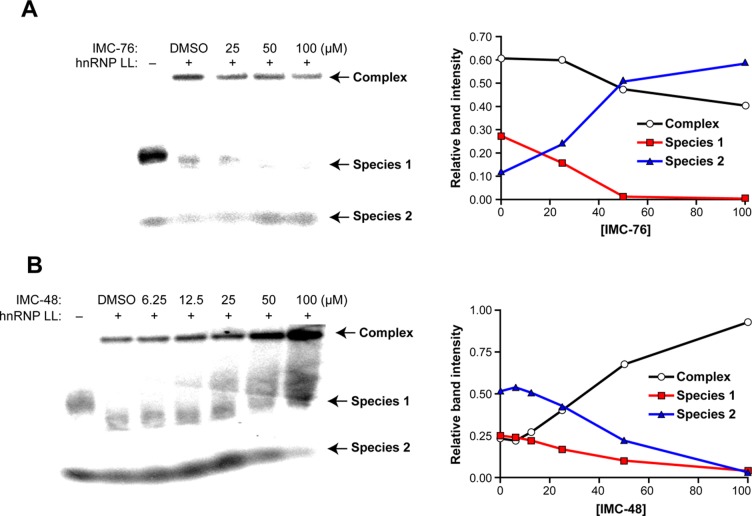
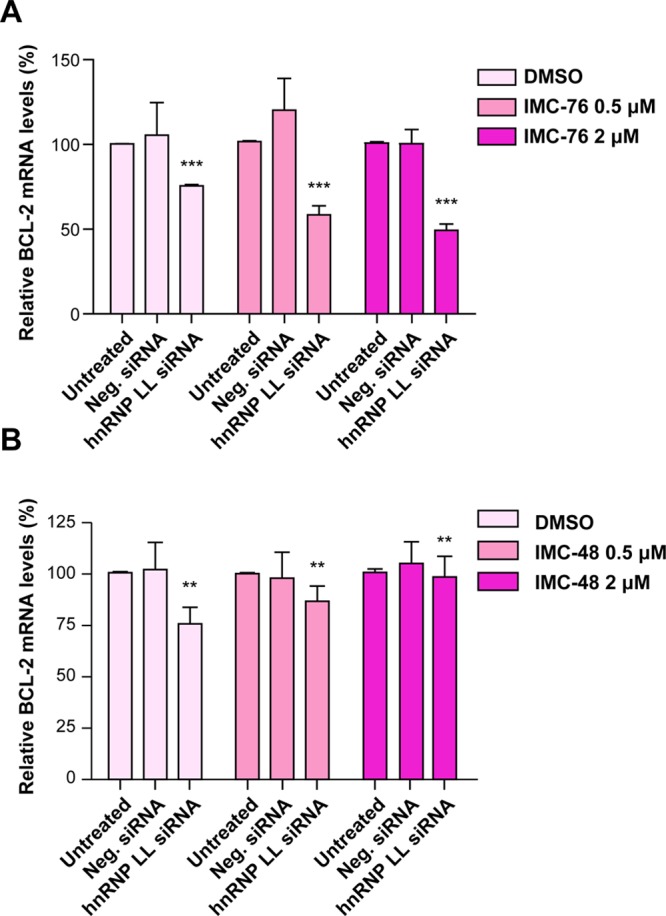
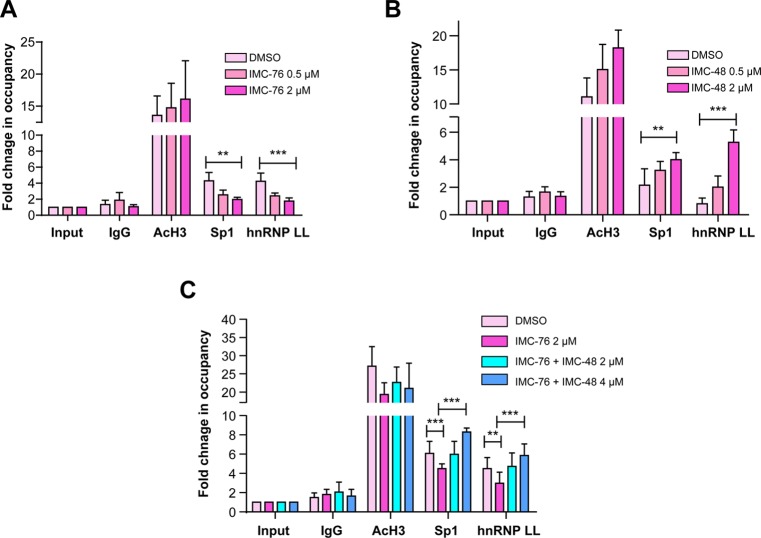
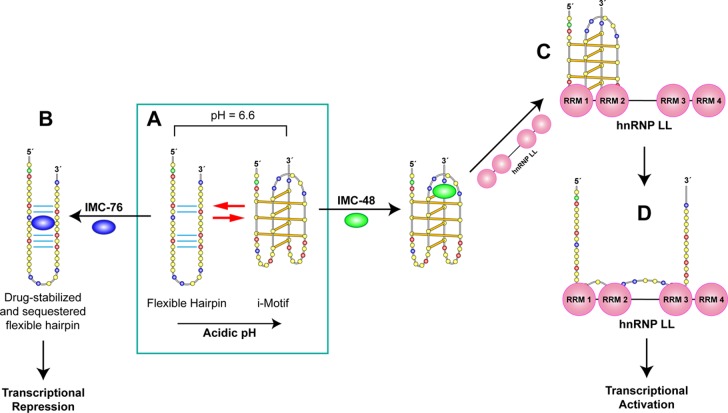
In a companion paper (DOI: 10.021/ja410934b) we demonstrate that the C-rich strand of the cis-regulatory element in the BCL2 promoter element is highly dynamic in nature and can form either an i-motif or a flexible hairpin. Under physiological conditions these two secondary DNA structures are found in an equilibrium mixture, which can be shifted by the addition of small molecules that trap out either the i-motif (IMC-48) or the flexible hairpin (IMC-76). In cellular experiments we demonstrate that the addition of these molecules has opposite effects on BCL2 gene expression and furthermore that these effects are antagonistic. In this contribution we have identified a transcriptional factor that recognizes and binds to the BCL2 i-motif to activate transcription. The molecular basis for the recognition of the i-motif by hnRNP LL is determined, and we demonstrate that the protein unfolds the i-motif structure to form a stable single-stranded complex. In subsequent experiments we show that IMC-48 and IMC-76 have opposite, antagonistic effects on the formation of the hnRNP LL-i-motif complex as well as on the transcription factor occupancy at the BCL2 promoter. For the first time we propose that the i-motif acts as a molecular switch that controls gene expression and that small molecules that target the dynamic equilibrium of the i-motif and the flexible hairpin can differentially modulate gene expression.
Figures
Similar articles
-
Molecular population dynamics of DNA structures in a bcl-2 promoter sequence is regulated by small molecules and the transcription factor hnRNP LL.Nucleic Acids Res. 2014 May;42(9):5755-64. doi: 10.1093/nar/gku185. Epub 2014 Mar 7. Nucleic Acids Res. 2014. PMID: 24609386 Free PMC article.
-
The dynamic character of the BCL2 promoter i-motif provides a mechanism for modulation of gene expression by compounds that bind selectively to the alternative DNA hairpin structure.J Am Chem Soc. 2014 Mar 19;136(11):4161-71. doi: 10.1021/ja410934b. Epub 2014 Mar 7. J Am Chem Soc. 2014. PMID: 24559410 Free PMC article.
-
Interaction of the N-Terminal Tandem Domains of hnRNP LL with the BCL2 Promoter i-Motif DNA Sequence.Chembiochem. 2017 Oct 18;18(20):2033-2044. doi: 10.1002/cbic.201700390. Epub 2017 Sep 8. Chembiochem. 2017. PMID: 28805284
-
Interaction of Individual Structural Domains of hnRNP LL with the BCL2 Promoter i-Motif DNA.J Am Chem Soc. 2016 Aug 31;138(34):10950-62. doi: 10.1021/jacs.6b05036. Epub 2016 Aug 17. J Am Chem Soc. 2016. PMID: 27483029
-
Critical role of hnRNP A1 in activating KRAS transcription in pancreatic cancer cells: A molecular mechanism involving G4 DNA.Biochim Biophys Acta Gen Subj. 2017 May;1861(5 Pt B):1389-1398. doi: 10.1016/j.bbagen.2016.11.031. Epub 2016 Nov 22. Biochim Biophys Acta Gen Subj. 2017. PMID: 27888145 Review.
Cited by
-
Non-canonical DNA structures: Diversity and disease association.Front Genet. 2022 Sep 5;13:959258. doi: 10.3389/fgene.2022.959258. eCollection 2022. Front Genet. 2022. PMID: 36134025 Free PMC article. Review.
-
Epigenetic modification of cytosines fine tunes the stability of i-motif DNA.Nucleic Acids Res. 2020 Jan 10;48(1):55-62. doi: 10.1093/nar/gkz1082. Nucleic Acids Res. 2020. PMID: 31777919 Free PMC article.
-
DNA i-motif formation at neutral pH is driven by kinetic partitioning.Nucleic Acids Res. 2023 Apr 11;51(6):2950-2962. doi: 10.1093/nar/gkad119. Nucleic Acids Res. 2023. PMID: 36912102 Free PMC article.
-
Interplay of Guanine Oxidation and G-Quadruplex Folding in Gene Promoters.J Am Chem Soc. 2020 Jan 22;142(3):1115-1136. doi: 10.1021/jacs.9b11050. Epub 2020 Jan 9. J Am Chem Soc. 2020. PMID: 31880930 Free PMC article.
-
Single-molecule analysis of i-motif within self-assembled DNA duplexes and nanocircles.Nucleic Acids Res. 2019 Aug 22;47(14):7199-7212. doi: 10.1093/nar/gkz565. Nucleic Acids Res. 2019. PMID: 31287873 Free PMC article.
References
-
- Brooks T. A.; Hurley L. H. Nat. Rev. Cancer 2009, 9, 849. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P30 CA023074/CA/NCI NIH HHS/United States
- GM085585-02S1/GM/NIGMS NIH HHS/United States
- R01 GM083117/GM/NIGMS NIH HHS/United States
- CA153821/CA/NCI NIH HHS/United States
- CA023074/CA/NCI NIH HHS/United States
- GM085585-01/GM/NIGMS NIH HHS/United States
- R01 CA153821/CA/NCI NIH HHS/United States
- R01 GM085585/GM/NIGMS NIH HHS/United States
- ES06694/ES/NIEHS NIH HHS/United States
- T32CA09213/CA/NCI NIH HHS/United States
- P30 ES006694/ES/NIEHS NIH HHS/United States
- GM083117/GM/NIGMS NIH HHS/United States
- T32 CA009213/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
