

ApoE and Aβ in Alzheimer's disease: accidental encounters or partners?
- PMID: 24559670
- PMCID: PMC3983361
- DOI: 10.1016/j.neuron.2014.01.045
ApoE and Aβ in Alzheimer's disease: accidental encounters or partners?
Abstract
Among the three human apolipoprotein E (apoE) isoforms, apoE4 increases the risk of Alzheimer's disease (AD). While transporting cholesterol is a primary function, apoE also regulates amyloid-β (Aβ) metabolism, aggregation, and deposition. Although earlier work suggests that different affinities of apoE isoforms to Aβ might account for their effects on Aβ clearance, recent studies indicate that apoE also competes with Aβ for cellular uptake through apoE receptors. Thus, several factors probably determine the variable effects apoE has on Aβ. In this Review, we examine biochemical, structural, and functional studies and propose testable models that address the complex mechanisms underlying apoE-Aβ interaction and how apoE4 may increase AD risk and also serve as a target pathway for therapy.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
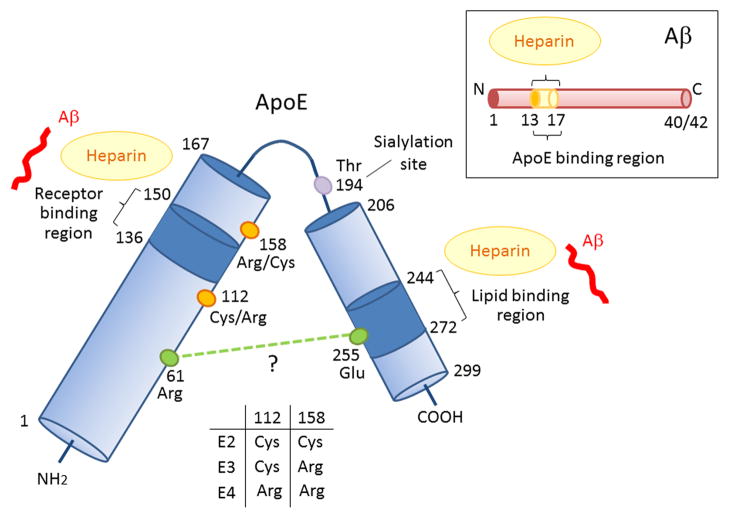

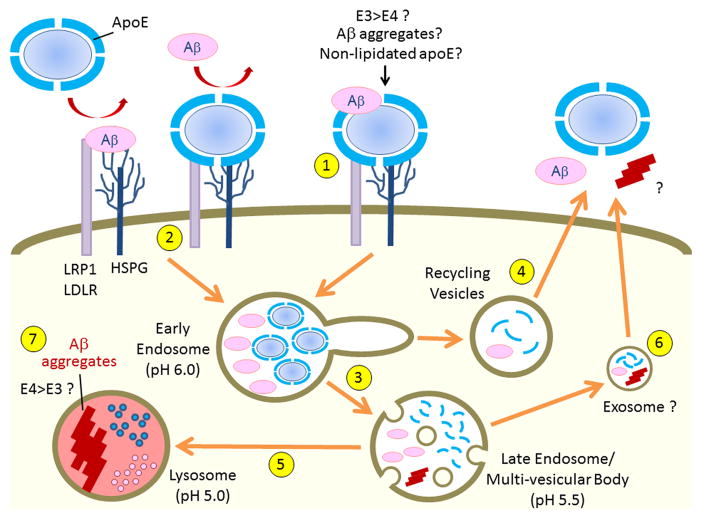
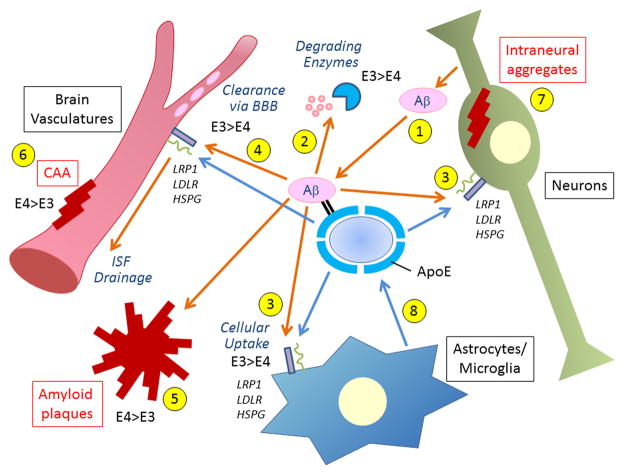
References
-
- Bales KR, Verina T, Dodel RC, Du Y, Altstiel L, Bender M, Hyslop P, Johnstone EM, Little SP, Cummins DJ, et al. Lack of apolipoprotein E dramatically reduces amyloid beta-peptide deposition. Nat Genet. 1997;17:263–264. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 AG035355/AG/NIA NIH HHS/United States
- R01 AG046205/AG/NIA NIH HHS/United States
- P01NS074969/NS/NINDS NIH HHS/United States
- R01 AG021173/AG/NIA NIH HHS/United States
- P01 NS074969/NS/NINDS NIH HHS/United States
- R01 AG044420/AG/NIA NIH HHS/United States
- R01AG046205/AG/NIA NIH HHS/United States
- R01AG027924/AG/NIA NIH HHS/United States
- R01 AG031784/AG/NIA NIH HHS/United States
- R01 AG027924/AG/NIA NIH HHS/United States
- P01 AG030128/AG/NIA NIH HHS/United States
- R01AG035355/AG/NIA NIH HHS/United States
- RF1 AG046205/AG/NIA NIH HHS/United States
- P01AG030128/AG/NIA NIH HHS/United States
- R01 NS046673/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
