

A local glutamate-glutamine cycle sustains synaptic excitatory transmitter release
- PMID: 24559677
- PMCID: PMC4001919
- DOI: 10.1016/j.neuron.2013.12.026
A local glutamate-glutamine cycle sustains synaptic excitatory transmitter release
Abstract
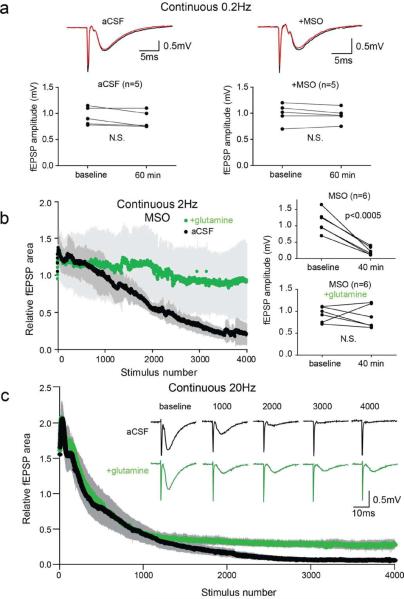
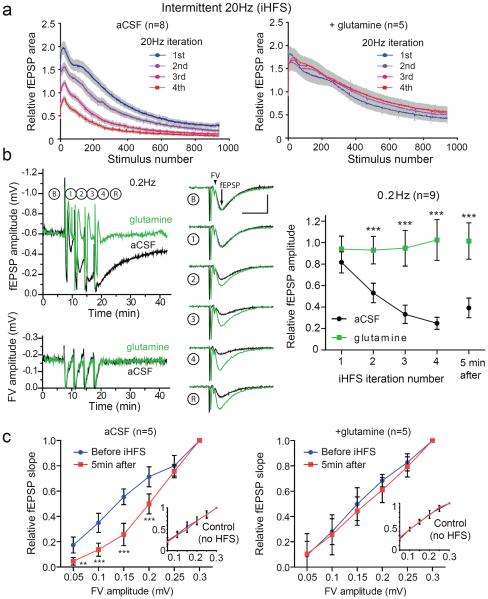
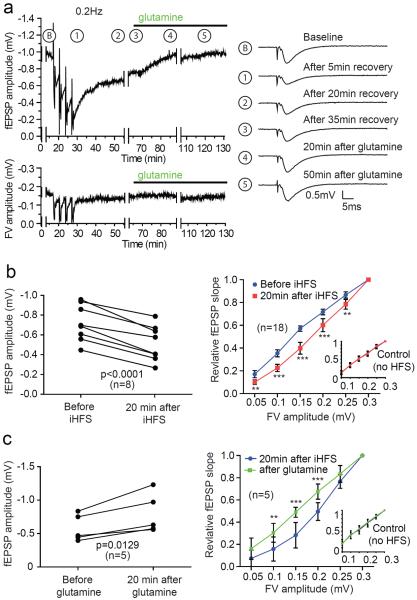
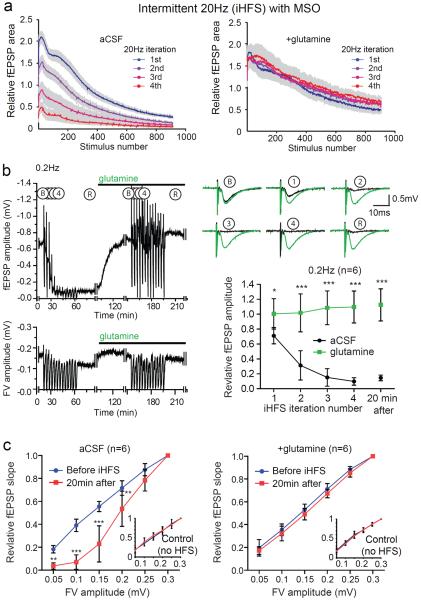
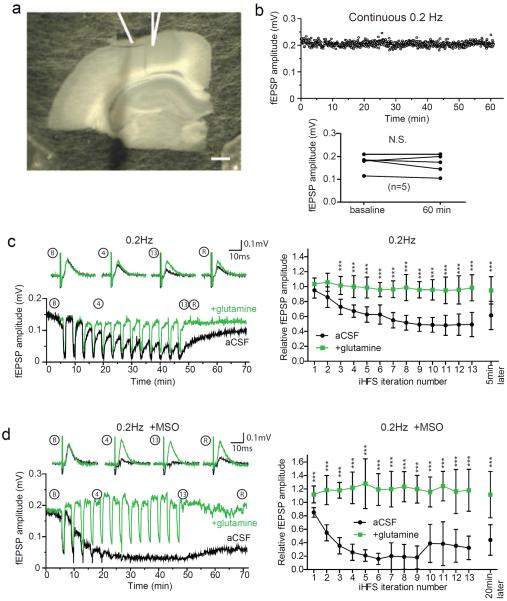
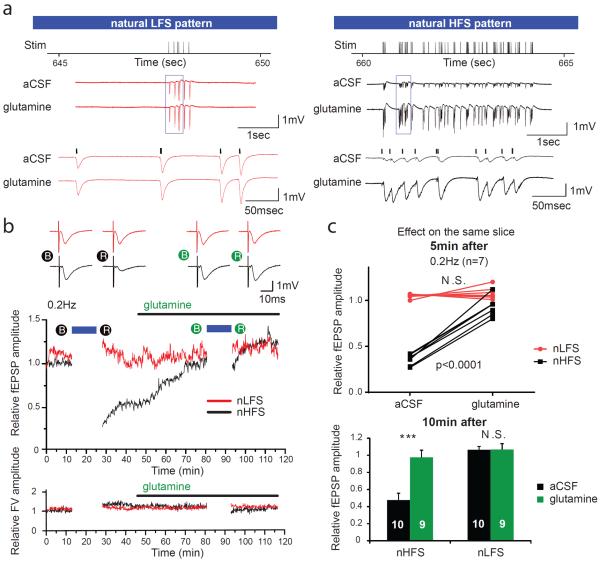
Biochemical studies suggest that excitatory neurons are metabolically coupled with astrocytes to generate glutamate for release. However, the extent to which glutamatergic neurotransmission depends on this process remains controversial because direct electrophysiological evidence is lacking. The distance between cell bodies and axon terminals predicts that glutamine-glutamate cycle is synaptically localized. Hence, we investigated isolated nerve terminals in brain slices by transecting hippocampal Schaffer collaterals and cortical layer I axons. Stimulating with alternating periods of high frequency (20 Hz) and rest (0.2 Hz), we identified an activity-dependent reduction in synaptic efficacy that correlated with reduced glutamate release. This was enhanced by inhibition of astrocytic glutamine synthetase and reversed or prevented by exogenous glutamine. Importantly, this activity dependence was also revealed with an in-vivo-derived natural stimulus both at network and cellular levels. These data provide direct electrophysiological evidence that an astrocyte-dependent glutamate-glutamine cycle is required to maintain active neurotransmission at excitatory terminals.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures


Comment in
-
Good housekeeping.Neuron. 2014 Feb 19;81(4):715-7. doi: 10.1016/j.neuron.2014.02.004. Neuron. 2014. PMID: 24559665 Free PMC article.
References
-
- Aoki C, Kaneko T, Starr A, Pickel VM. Identification of mitochondrial and non-mitochondrial glutaminase within select neurons and glia of rat forebrain by electron microscopic immunocytochemistry. J Neurosci Res. 1991;28:531–548. - PubMed
-
- Bacci A, Sancini G, Verderio C, Armano S, Pravettoni E, Fesce R, Franceschetti S, Matteoli M. Block of glutamate-glutamine cycle between astrocytes and neurons inhibits epileptiform activity in hippocampus. J Neurophysiol. 2002;88:2302–2310. - PubMed
-
- Balkrishna S, Broer A, Kingsland A, Broer S. Rapid downregulation of the rat glutamine transporter SNAT3 by a caveolin-dependent trafficking mechanism in Xenopus laevis oocytes. Am J Physiol Cell Physiol. 2010;299:C1047–1057. - PubMed
-
- Berke JD, Hetrick V, Breck J, Greene RW. Transient 23–30 Hz oscillations in mouse hippocampus during exploration of novel environments. Hippocampus. 2008;18:519–529. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
