

Dopamine neurons control striatal cholinergic neurons via regionally heterogeneous dopamine and glutamate signaling
- PMID: 24559678
- PMCID: PMC3933825
- DOI: 10.1016/j.neuron.2013.12.027
Dopamine neurons control striatal cholinergic neurons via regionally heterogeneous dopamine and glutamate signaling
Abstract
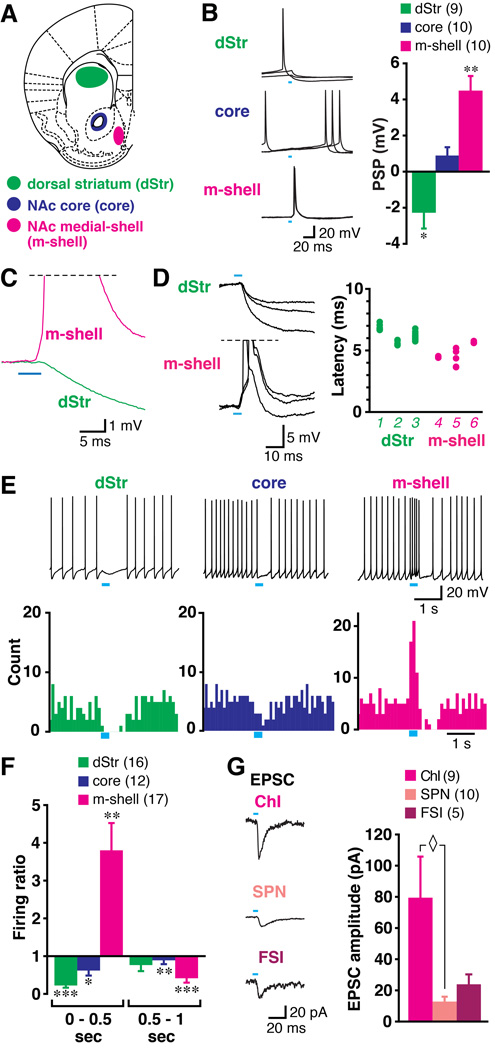
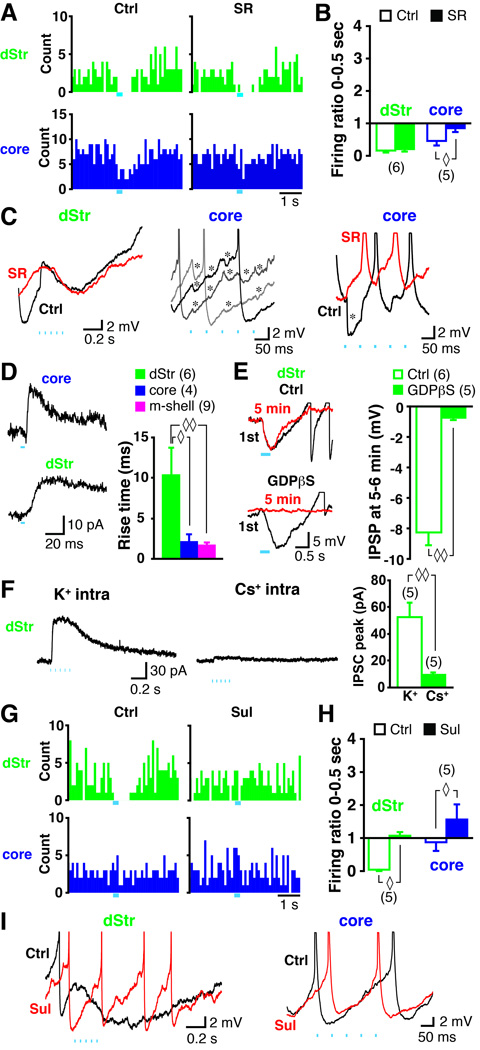
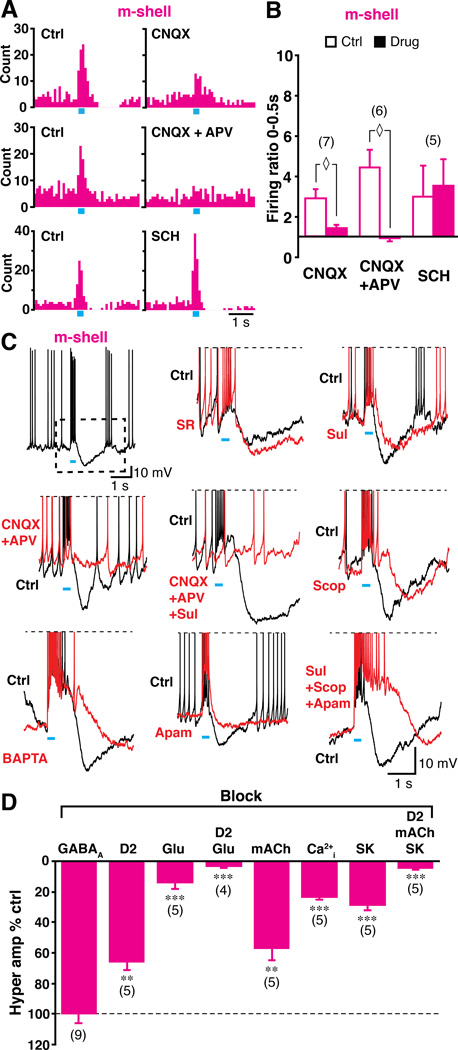
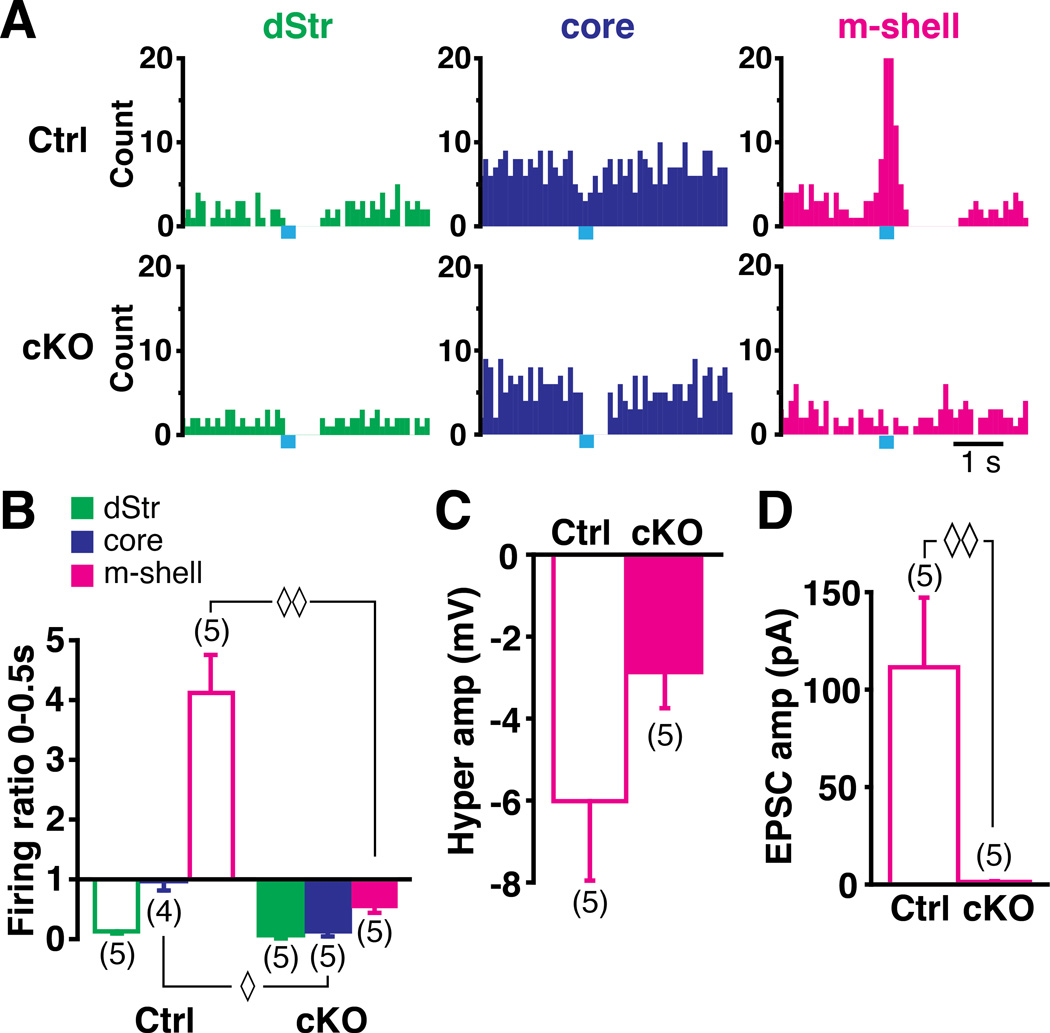
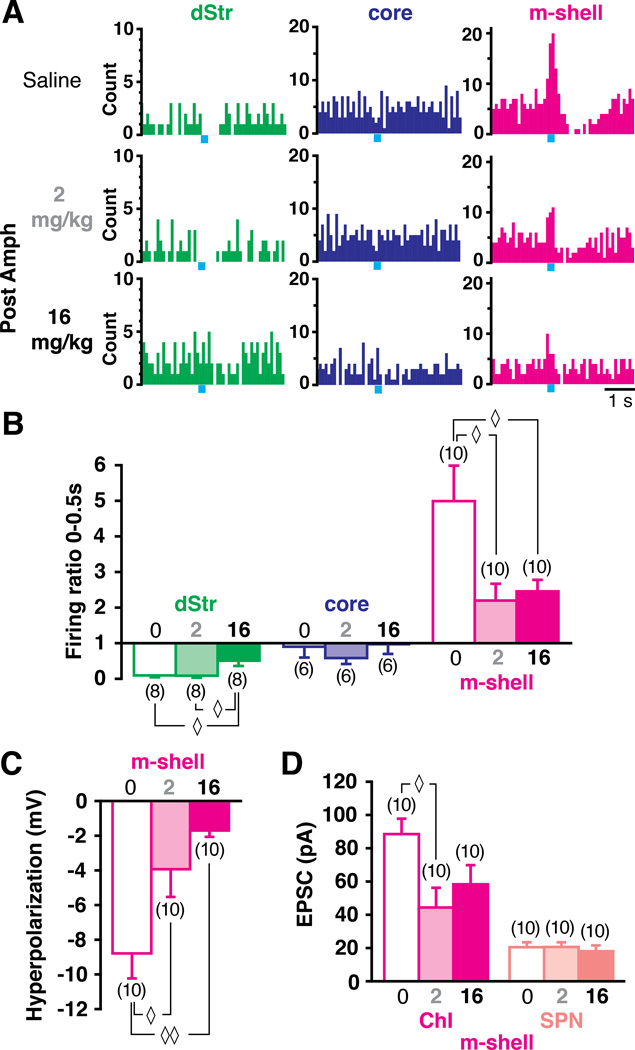
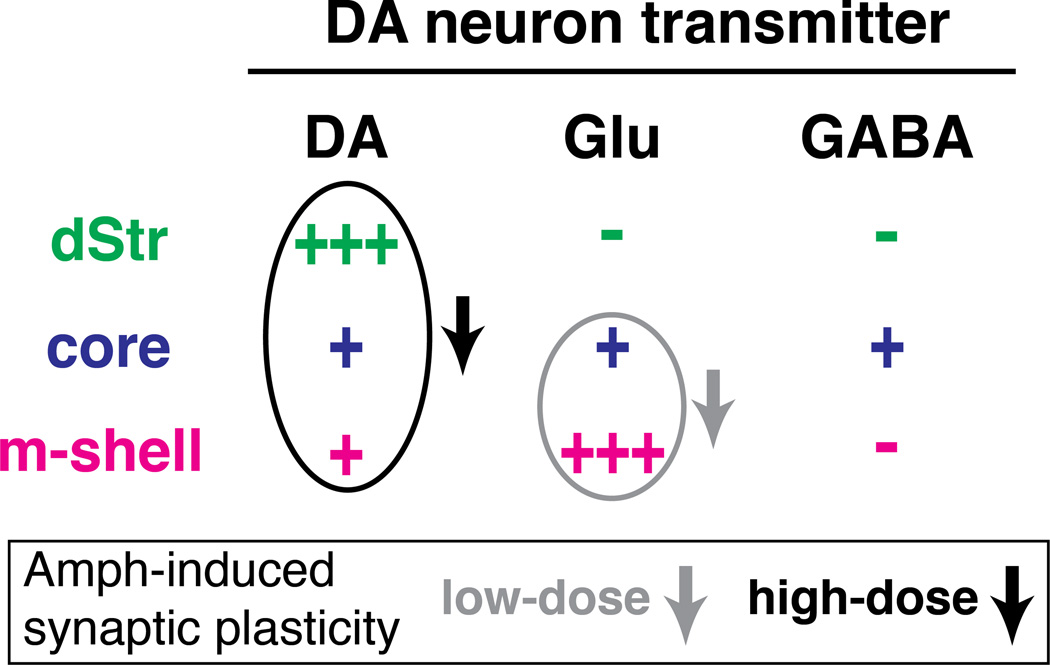
Midbrain dopamine neurons fire in bursts conveying salient information. Bursts are associated with pauses in tonic firing of striatal cholinergic interneurons. Although the reciprocal balance of dopamine and acetylcholine in the striatum is well known, how dopamine neurons control cholinergic neurons has not been elucidated. Here, we show that dopamine neurons make direct fast dopaminergic and glutamatergic connections with cholinergic interneurons, with regional heterogeneity. Dopamine neurons drive a burst-pause firing sequence in cholinergic interneurons in the medial shell of the nucleus accumbens, mixed actions in the accumbens core, and a pause in the dorsal striatum. This heterogeneity is due mainly to regional variation in dopamine-neuron glutamate cotransmission. A single dose of amphetamine attenuates dopamine neuron connections to cholinergic interneurons with dose-dependent regional specificity. Overall, the present data indicate that dopamine neurons control striatal circuit function via discrete, plastic connections with cholinergic interneurons.
Copyright © 2014 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors have no conflicts of interest to declare.
Figures

References
-
- Bäckman CM, Malik N, Zhang Y, Shan L, Grinberg A, Hoffer BJ, Westphal H, Tomac AC. Characterization of a mouse strain expressing Cre recombinase from the 3' untranslated region of the dopamine transporter locus. Genesis. 2006;44:383–390. - PubMed
-
- Beckstead MJ, Grandy DK, Wickman K, Williams JT. Vesicular dopamine release elicits an inhibitory postsynaptic current in midbrain dopamine neurons. Neuron. 2004;42:939–946. - PubMed
-
- Belin D, Jonkman S, Dickinson A, Robbins TW, Everitt BJ. Parallel and interactive learning processes within the basal ganglia: relevance for the understanding of addiction. Behav Brain Res. 2009;199:89–102. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
