

Nitric oxide-induced conformational changes in soluble guanylate cyclase
- PMID: 24560804
- PMCID: PMC4001857
- DOI: 10.1016/j.str.2014.01.008
Nitric oxide-induced conformational changes in soluble guanylate cyclase
Abstract
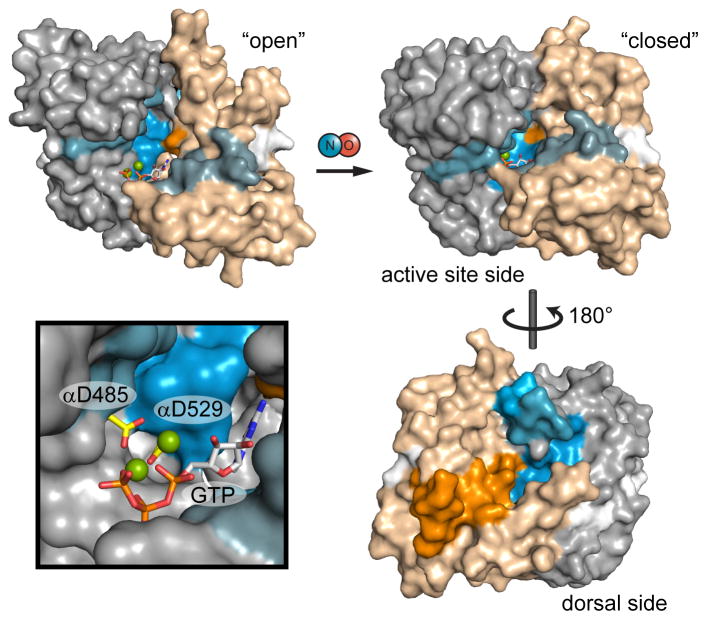
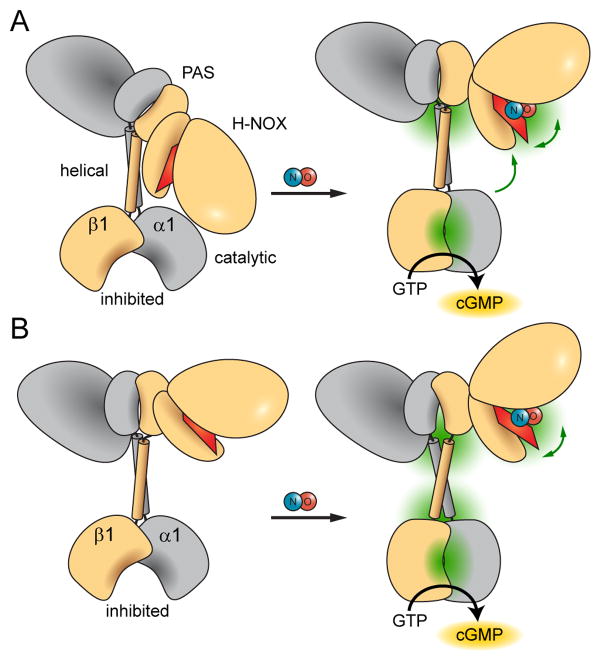
Soluble guanylate cyclase (sGC) is the primary mediator of nitric oxide (NO) signaling. NO binds the sGC heme cofactor stimulating synthesis of the second messenger cyclic-GMP (cGMP). As the central hub of NO/cGMP signaling pathways, sGC is important in diverse physiological processes such as vasodilation and neurotransmission. Nevertheless, the mechanisms underlying NO-induced cyclase activation in sGC remain unclear. Here, hydrogen/deuterium exchange mass spectrometry (HDX-MS) was employed to probe the NO-induced conformational changes of sGC. HDX-MS revealed NO-induced effects in several discrete regions. NO binding to the heme-NO/O2-binding (H-NOX) domain perturbs a signaling surface implicated in Per/Arnt/Sim (PAS) domain interactions. Furthermore, NO elicits striking conformational changes in the junction between the PAS and helical domains that propagate as perturbations throughout the adjoining helices. Ultimately, NO binding stimulates the catalytic domain by contracting the active site pocket. Together, these conformational changes delineate an allosteric pathway linking NO binding to activation of the catalytic domain.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures
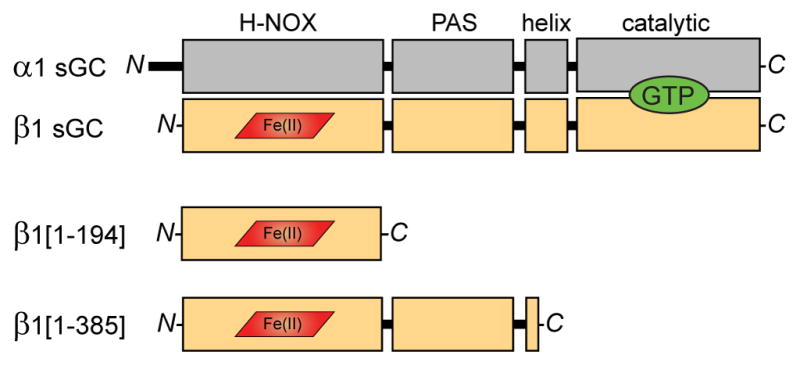
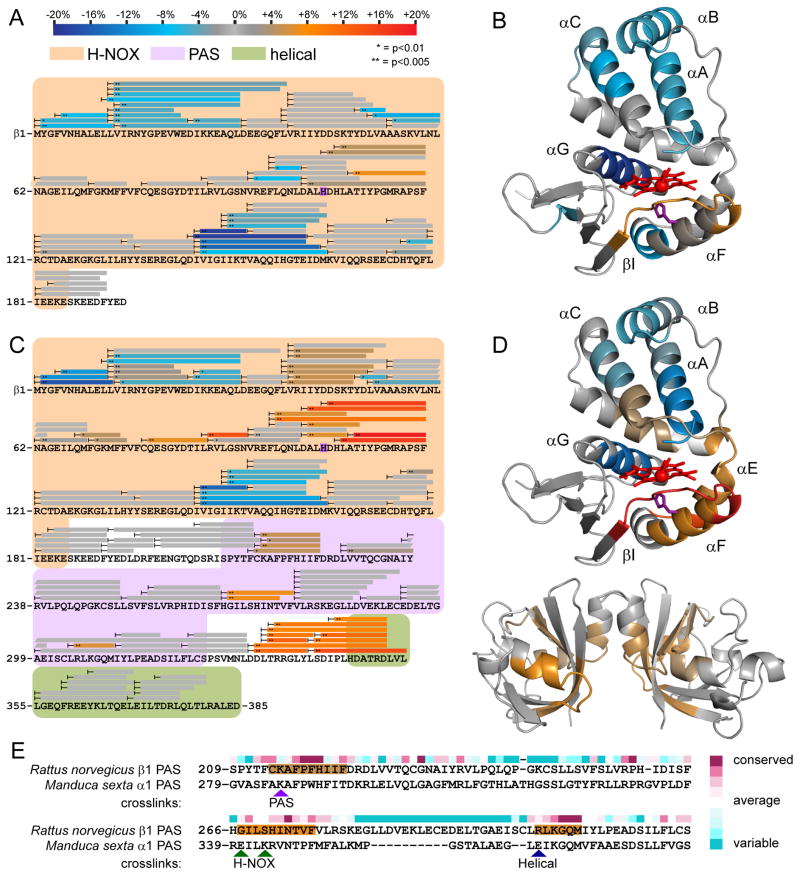
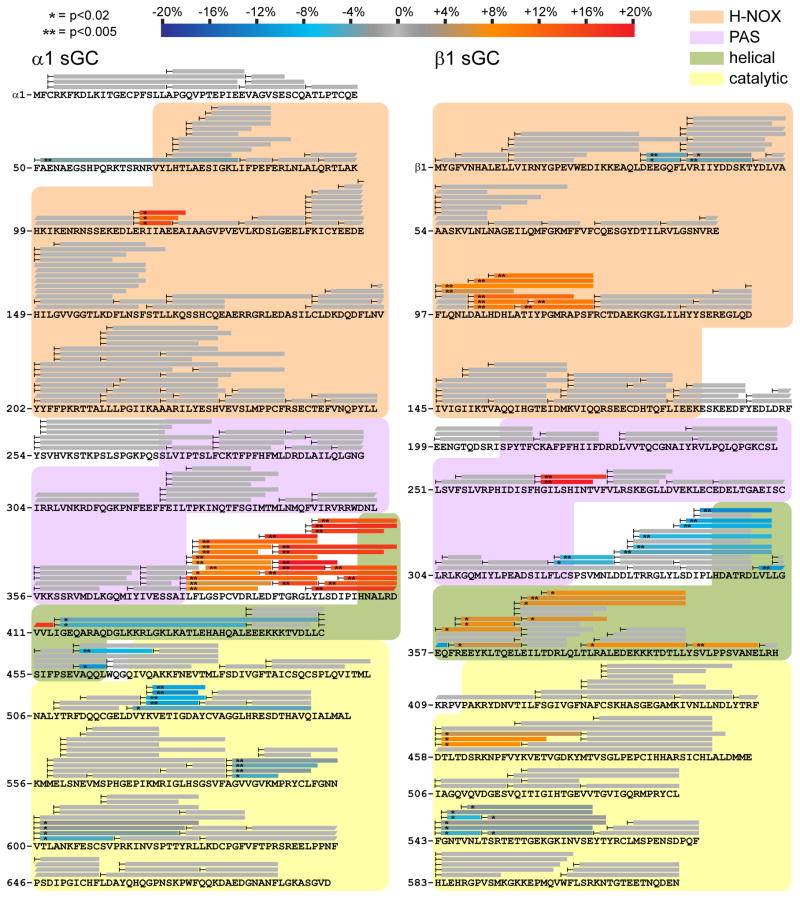
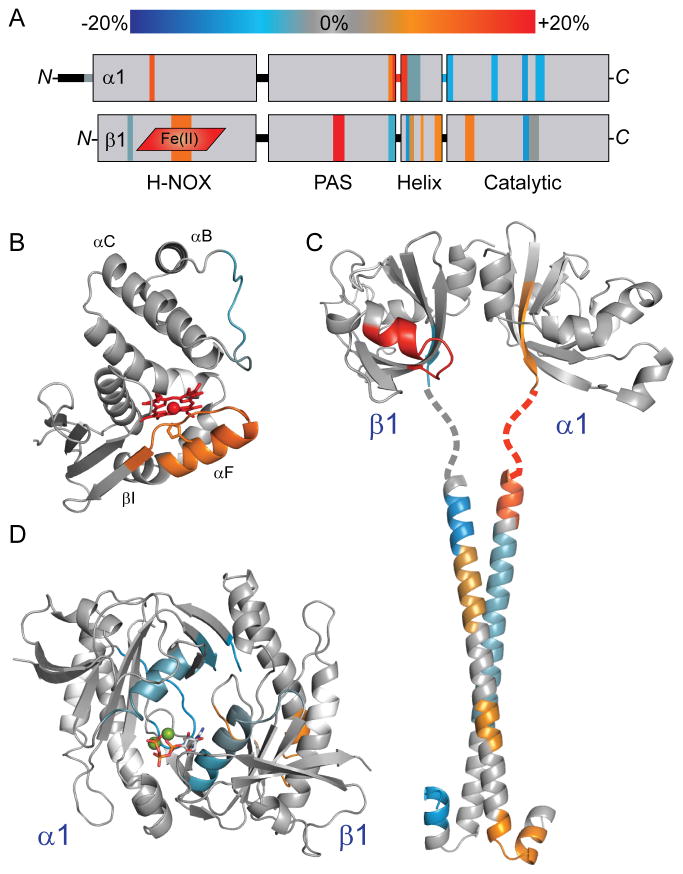
Comment in
-
Tracking allosteric propagation with HX-MS.Structure. 2014 Apr 8;22(4):512-4. doi: 10.1016/j.str.2014.03.005. Structure. 2014. PMID: 24717559
References
-
- Bredt DS. Endogenous nitric oxide synthesis: Biological functions and pathophysiology. Free Radic Res. 1999;31:577–596. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
