

Polyphosphate is a primordial chaperone
- PMID: 24560923
- PMCID: PMC3996911
- DOI: 10.1016/j.molcel.2014.01.012
Polyphosphate is a primordial chaperone
Abstract
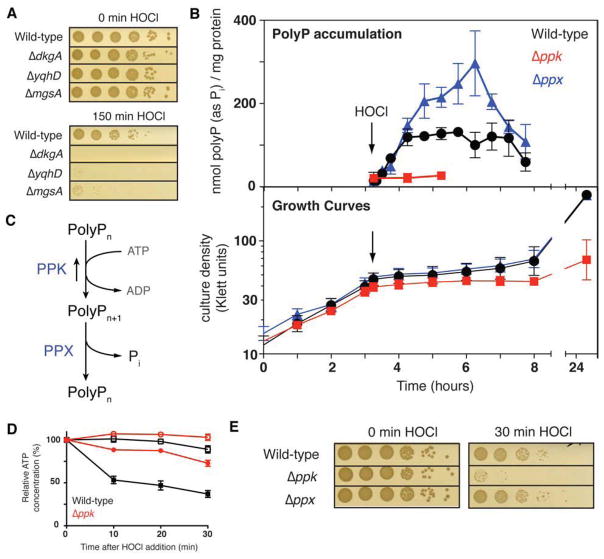
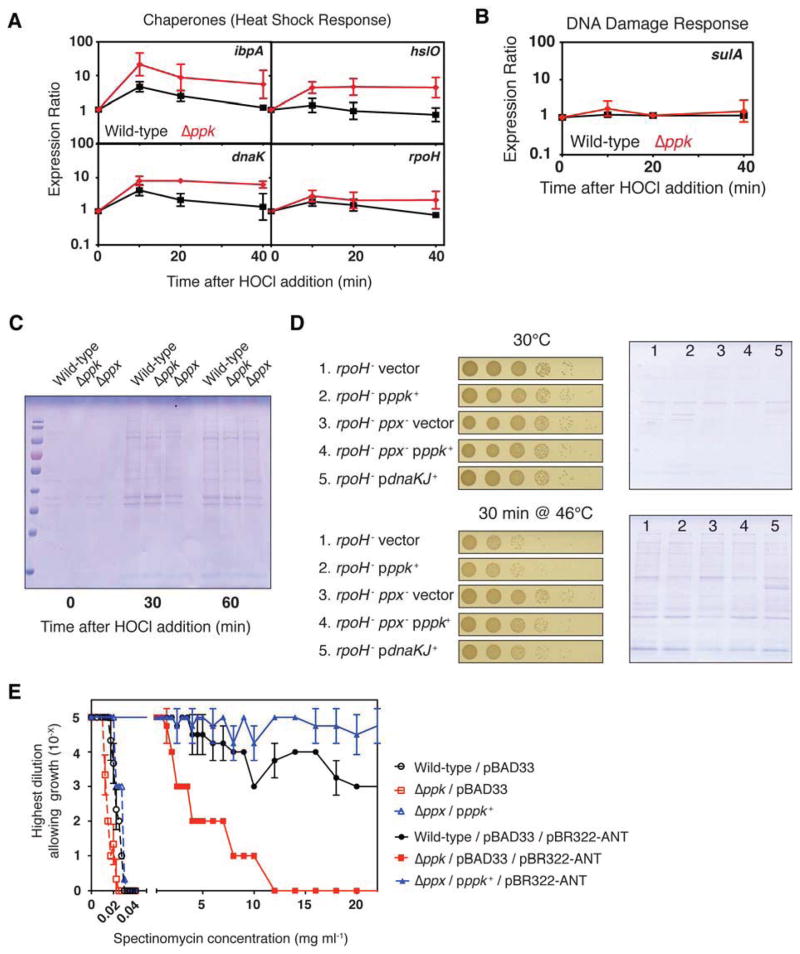
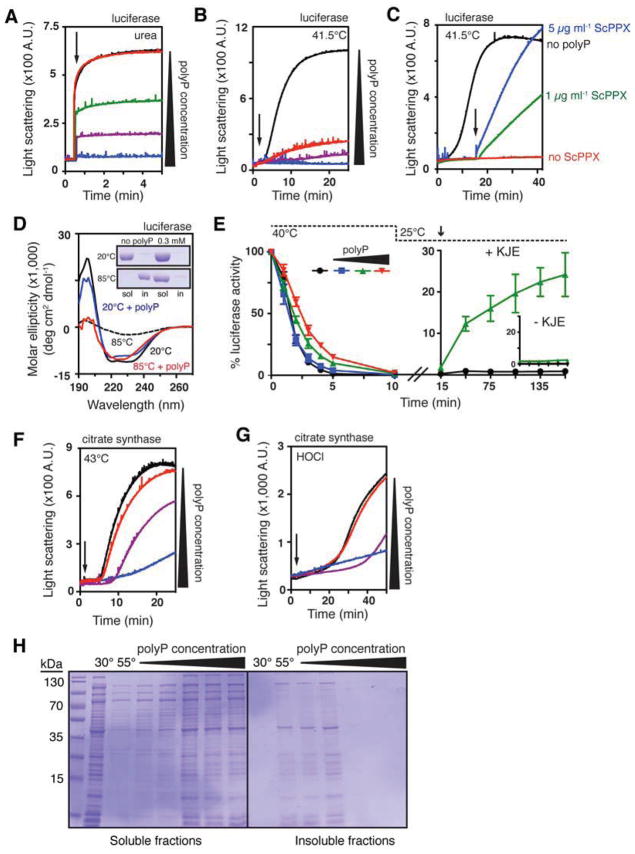
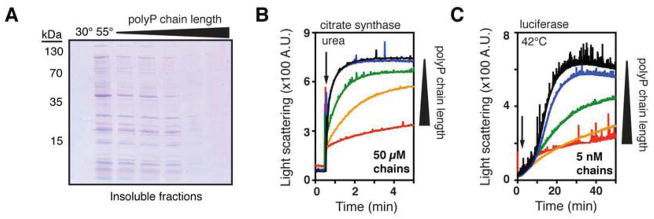
Composed of up to 1,000 phospho-anhydride bond-linked phosphate monomers, inorganic polyphosphate (polyP) is one of the most ancient, conserved, and enigmatic molecules in biology. Here we demonstrate that polyP functions as a hitherto unrecognized chaperone. We show that polyP stabilizes proteins in vivo, diminishes the need for other chaperone systems to survive proteotoxic stress conditions, and protects a wide variety of proteins against stress-induced unfolding and aggregation. In vitro studies reveal that polyP has protein-like chaperone qualities, binds to unfolding proteins with high affinity in an ATP-independent manner, and supports their productive refolding once nonstress conditions are restored. Our results uncover a universally important function for polyP and suggest that these long chains of inorganic phosphate may have served as one of nature's first chaperones, a role that continues to the present day.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
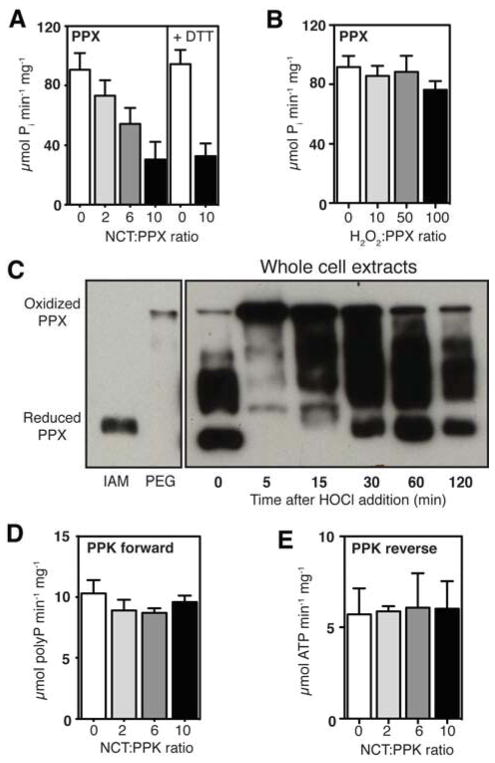
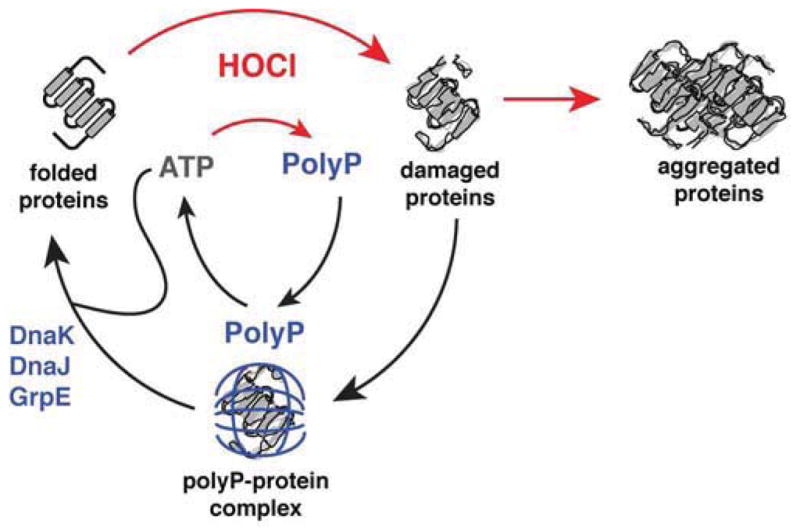
Comment in
-
Chaperoned by prebiotic inorganic polyphosphate molecules: an ancient transcription-independent mechanism to restore protein homeostasis.Mol Cell. 2014 Mar 6;53(5):685-7. doi: 10.1016/j.molcel.2014.02.023. Mol Cell. 2014. PMID: 24606917
References
-
- Alvarado J, Ghosh A, Janovitz T, Jauregui A, Hasson MS, Sanders DA. Origin of exopolyphosphatase processivity: Fusion of an ASKHA phosphotransferase and a cyclic nucleotide phosphodiesterase homolog. Structure. 2006;14:1263–1272. - PubMed
-
- Anfinsen CB. Principles that govern the folding of protein chains. Science. 1973;181:223–230. - PubMed
-
- Aschar-Sobbi R, Abramov AY, Diao C, Kargacin ME, Kargacin GJ, French RJ, Pavlov E. High sensitivity, quantitative measurements of polyphosphate using a new DAPI-based approach. J Fluoresc. 2008;18:859–866. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
