

Role of tryptophan side chain dynamics on the Trp-cage mini-protein folding studied by molecular dynamics simulations
- PMID: 24563686
- PMCID: PMC3921895
- DOI: 10.1371/journal.pone.0088383
Role of tryptophan side chain dynamics on the Trp-cage mini-protein folding studied by molecular dynamics simulations
Abstract
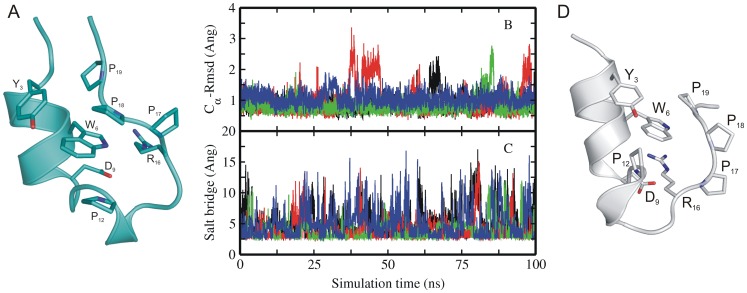
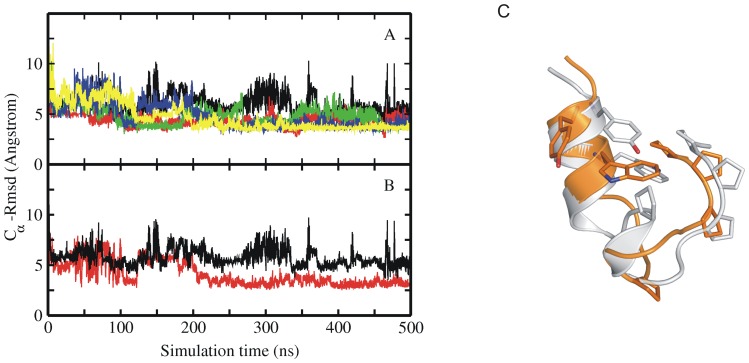
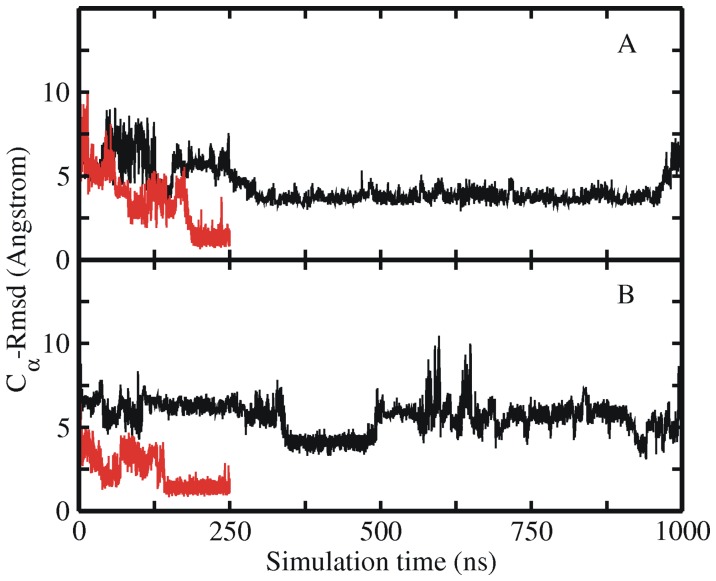
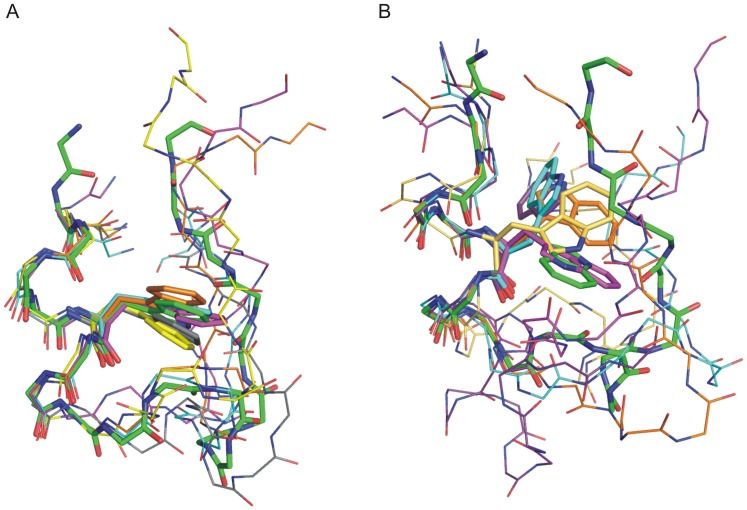
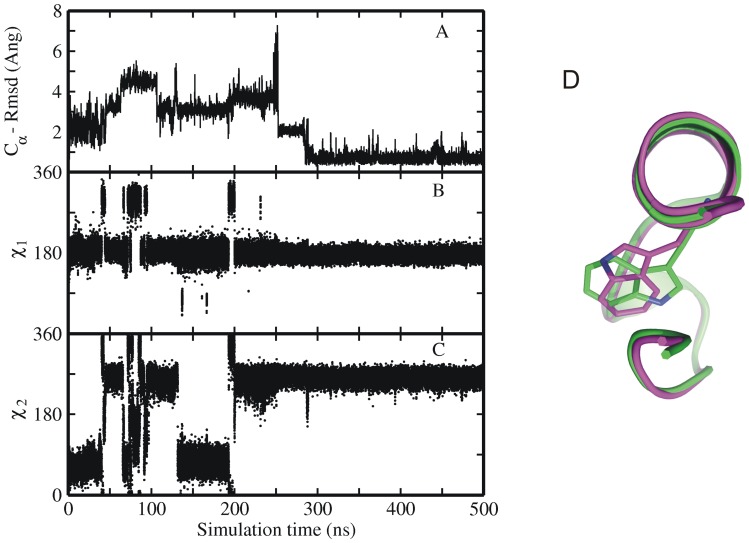
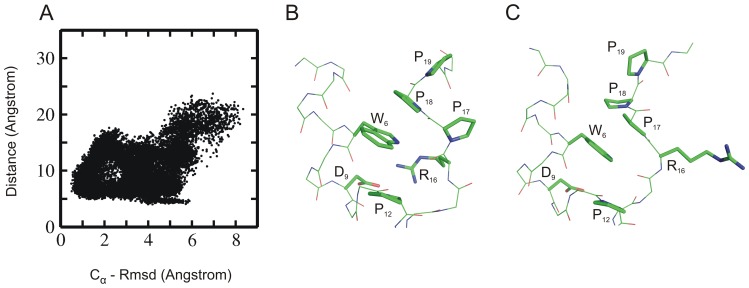
The 20 residue Trp-cage mini-protein is one of smallest proteins that adopt a stable folded structure containing also well-defined secondary structure elements. The hydrophobic core is arranged around a single central Trp residue. Despite several experimental and simulation studies the detailed folding mechanism of the Trp-cage protein is still not completely understood. Starting from fully extended as well as from partially folded Trp-cage structures a series of molecular dynamics simulations in explicit solvent and using four different force fields was performed. All simulations resulted in rapid collapse of the protein to on average relatively compact states. The simulations indicate a significant dependence of the speed of folding to near-native states on the side chain rotamer state of the central Trp residue. Whereas the majority of intermediate start structures with the central Trp side chain in a near-native rotameric state folded successfully within less than 100 ns only a fraction of start structures reached near-native folded states with an initially non-native Trp side chain rotamer state. Weak restraining of the Trp side chain dihedral angles to the state in the folded protein resulted in significant acceleration of the folding both starting from fully extended or intermediate conformations. The results indicate that the side chain conformation of the central Trp residue can create a significant barrier for controlling transitions to a near native folded structure. Similar mechanisms might be of importance for the folding of other protein structures.
Conflict of interest statement
Figures
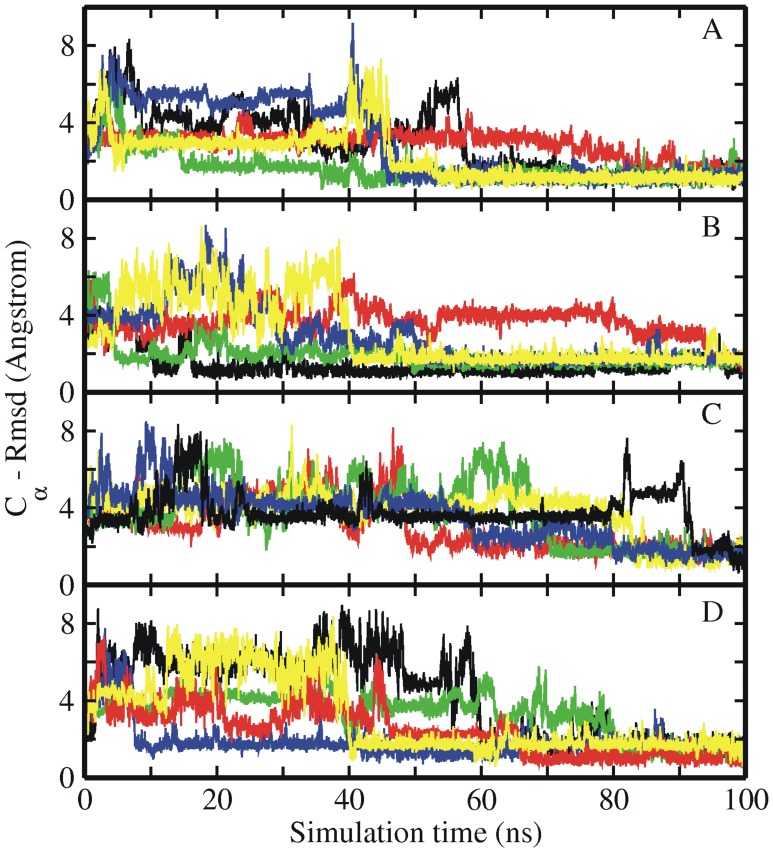
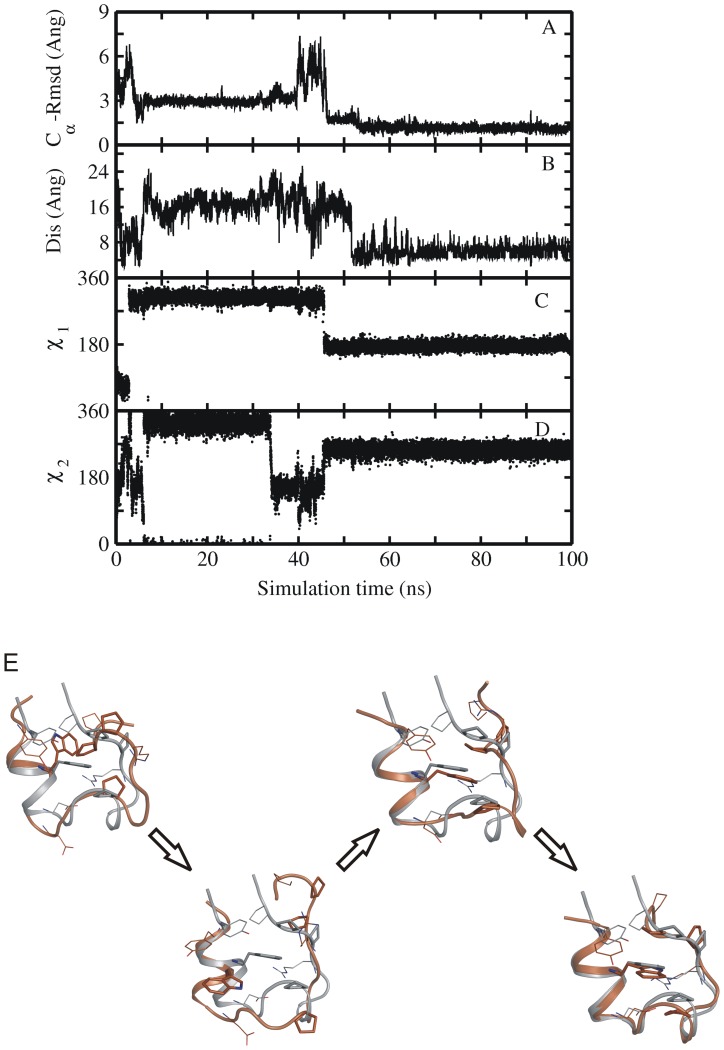
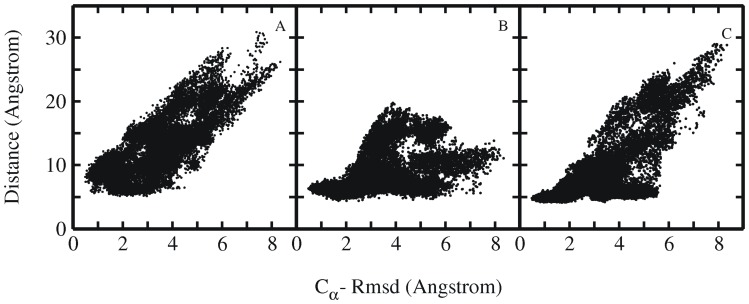
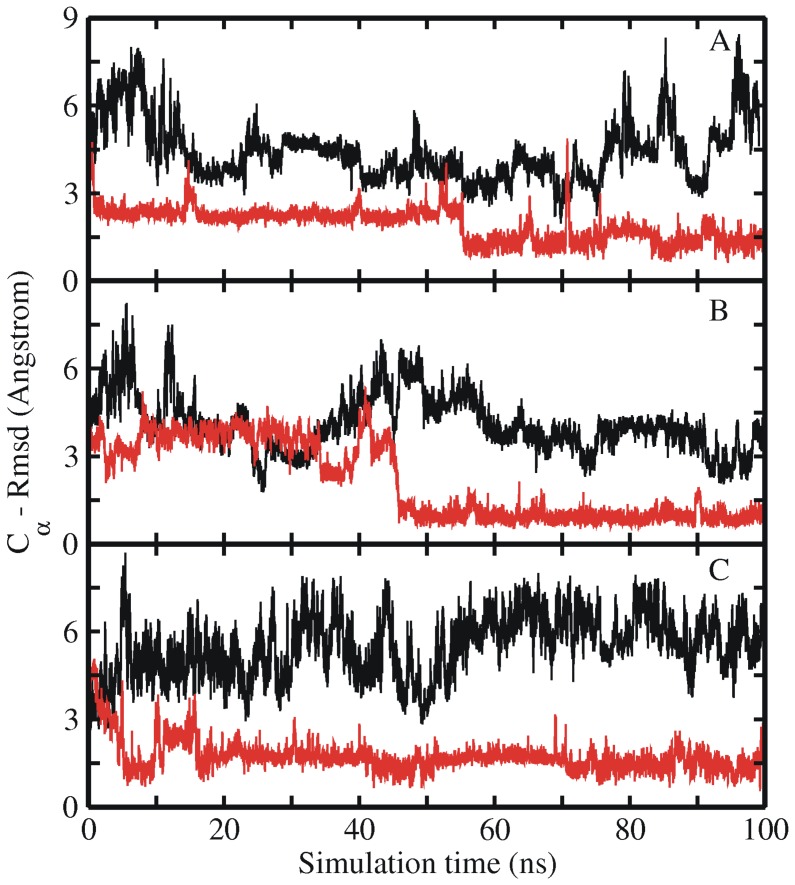
References
-
- Neidigh JW, Fesinmeyer MR, Andersen HN (2002) Designing a 20-residue protein. Nat. Struct. Biol. 9: 425–430. - PubMed
-
- Qiu L, Pabit SA, Roitberg AE, Hagen SJ (2002) Smaller and faster: the 20-residue Trp-cage protein folds in 4 micros. J. Am. Chem. Soc. 124: 12952–12953. - PubMed
-
- Streicher WW, Makhatadze GI (2007) Unfolding thermodynamics of Trp-cage, a 20 residue miniprotein, studied by differential scanning calorimetry and circular dichroism spectroscopy. Biochemistry. 46: 2876–2880. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
