

Mycobacterium abscessus cording prevents phagocytosis and promotes abscess formation
- PMID: 24567393
- PMCID: PMC3956181
- DOI: 10.1073/pnas.1321390111
Mycobacterium abscessus cording prevents phagocytosis and promotes abscess formation
Abstract
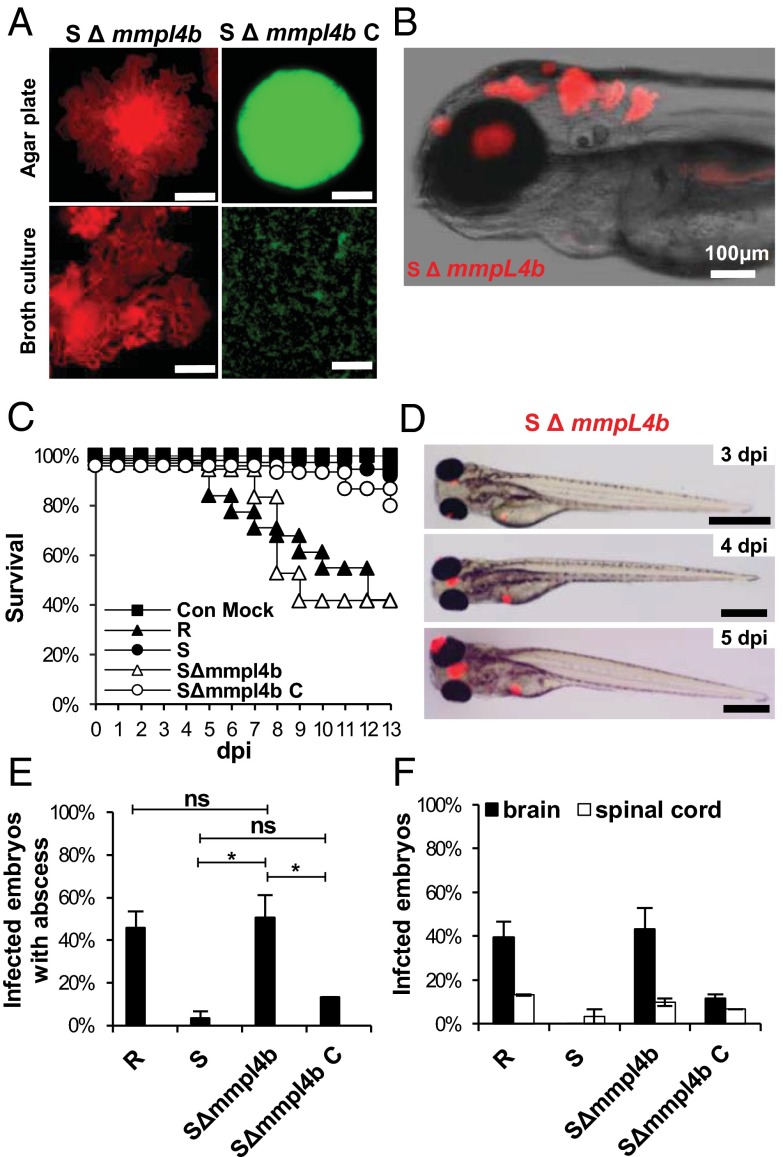
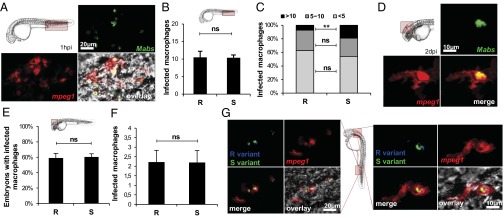
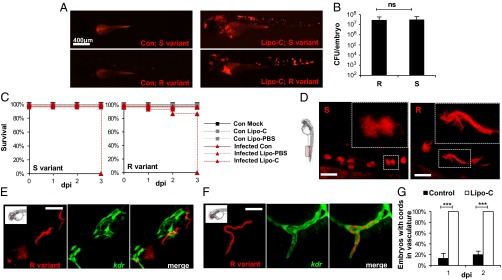
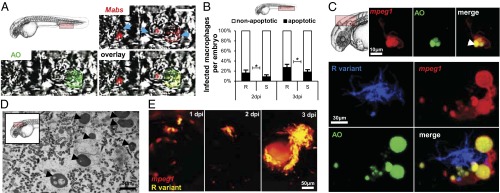
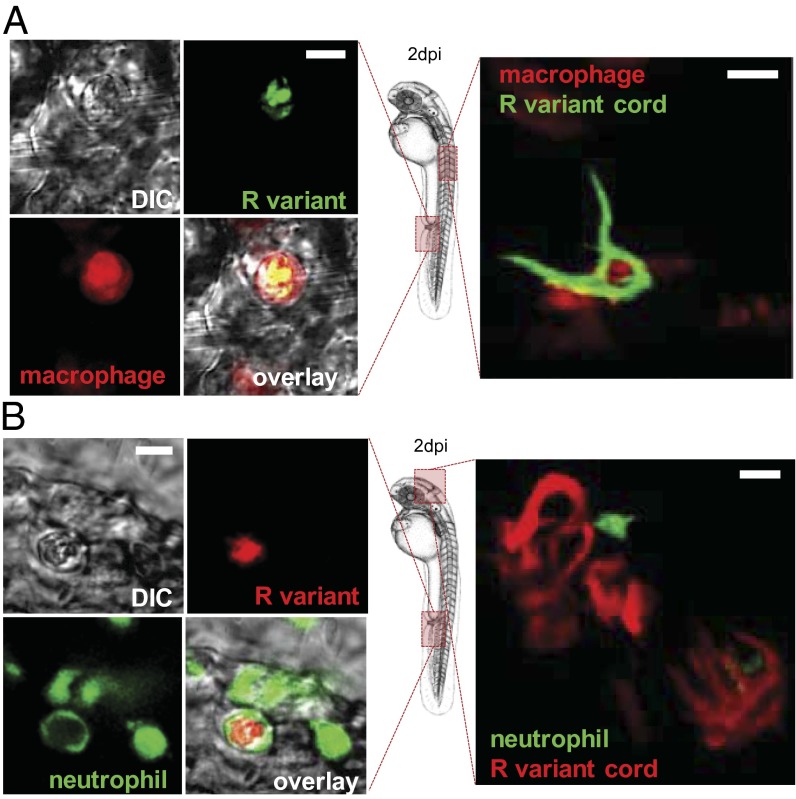
Mycobacterium abscessus is a rapidly growing Mycobacterium causing a wide spectrum of clinical syndromes. It now is recognized as a pulmonary pathogen to which cystic fibrosis patients have a particular susceptibility. The M. abscessus rough (R) variant, devoid of cell-surface glycopeptidolipids (GPLs), causes more severe clinical disease than the smooth (S) variant, but the underlying mechanisms of R-variant virulence remain obscure. Exploiting the optical transparency of zebrafish embryos, we observed that the increased virulence of the M. abscessus R variant compared with the S variant correlated with the loss of GPL production. The virulence of the R variant involved the massive production of serpentine cords, absent during S-variant infection, and the cords initiated abscess formation leading to rapid larval death. Cording occurred within the vasculature and was highly pronounced in the central nervous system (CNS). It appears that M. abscessus is transported to the CNS within macrophages. The release of M. abscessus from apoptotic macrophages initiated the formation of cords that grew too large to be phagocytized by macrophages or neutrophils. This study is a description of the crucial role of cording in the in vivo physiopathology of M. abscessus infection and emphasizes cording as a mechanism of immune evasion.
Keywords: granuloma; innate immunity; morphotype; pathogenesis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Wallace RG, Jr, Silcox V, Brown BA. Taxonomy of rapidly growing mycobacteria. Clin Infect Dis. 1994;18(1):121–122. - PubMed
-
- Olivier KN, et al. Nontuberculous Mycobacteria in Cystic Fibrosis Study Group Nontuberculous mycobacteria. I: Multicenter prevalence study in cystic fibrosis. Am J Respir Crit Care Med. 2003;167(6):828–834. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
