

CELLULOSE SYNTHASE-LIKE A2, a glucomannan synthase, is involved in maintaining adherent mucilage structure in Arabidopsis seed
- PMID: 24569843
- PMCID: PMC3982747
- DOI: 10.1104/pp.114.236596
CELLULOSE SYNTHASE-LIKE A2, a glucomannan synthase, is involved in maintaining adherent mucilage structure in Arabidopsis seed
Abstract
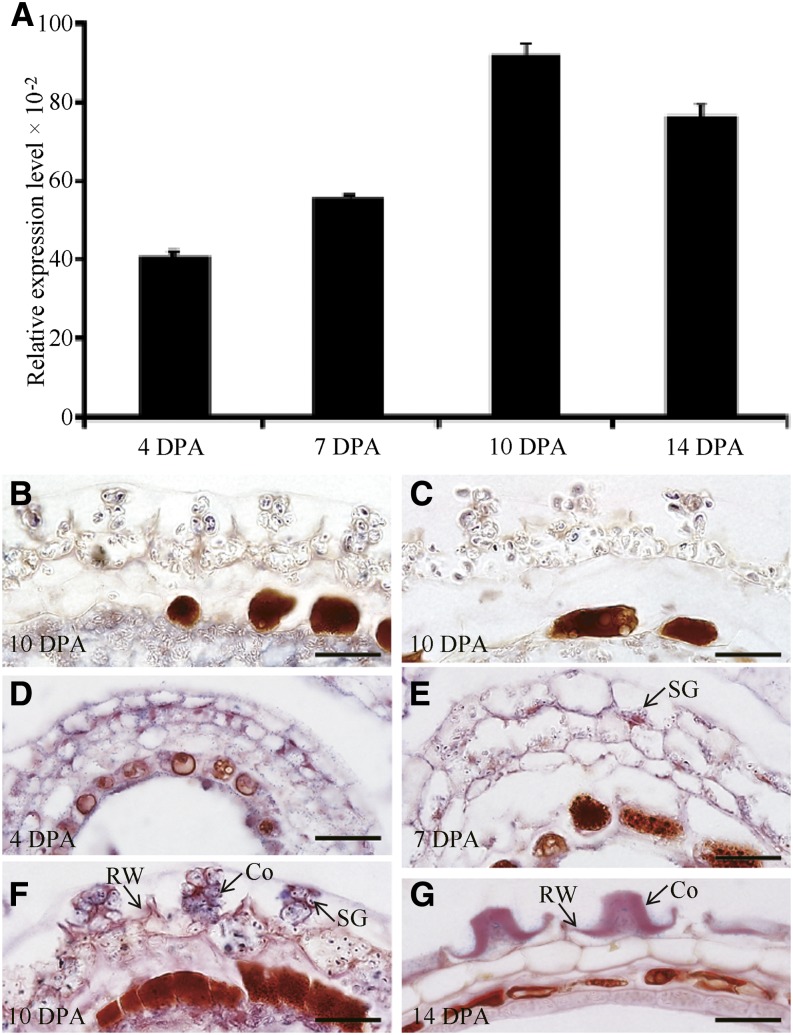
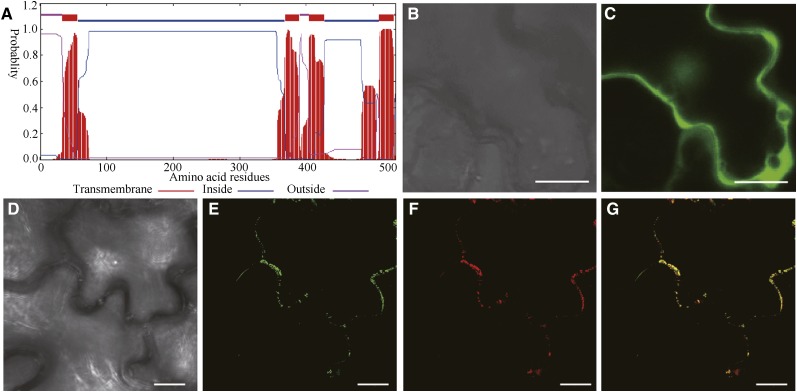
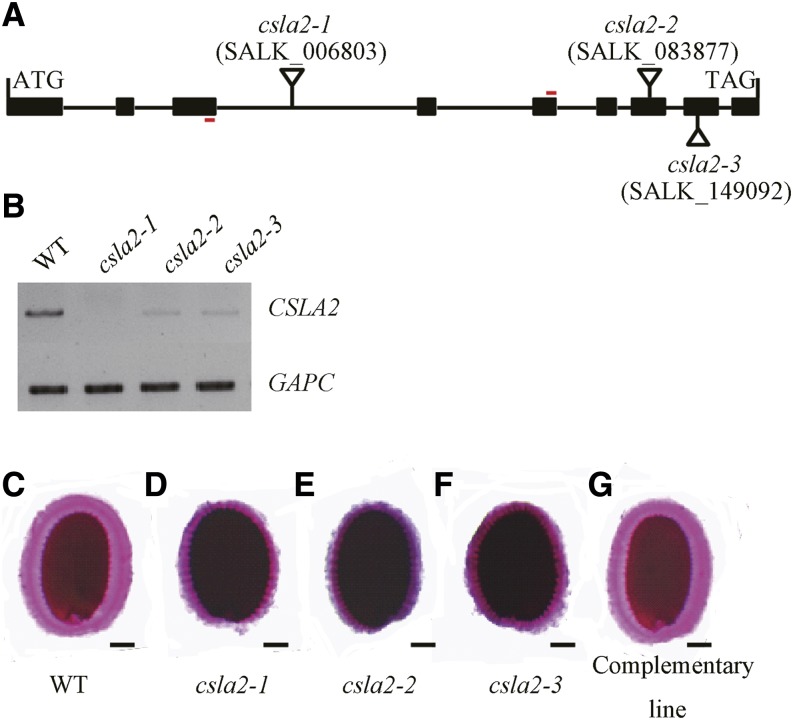
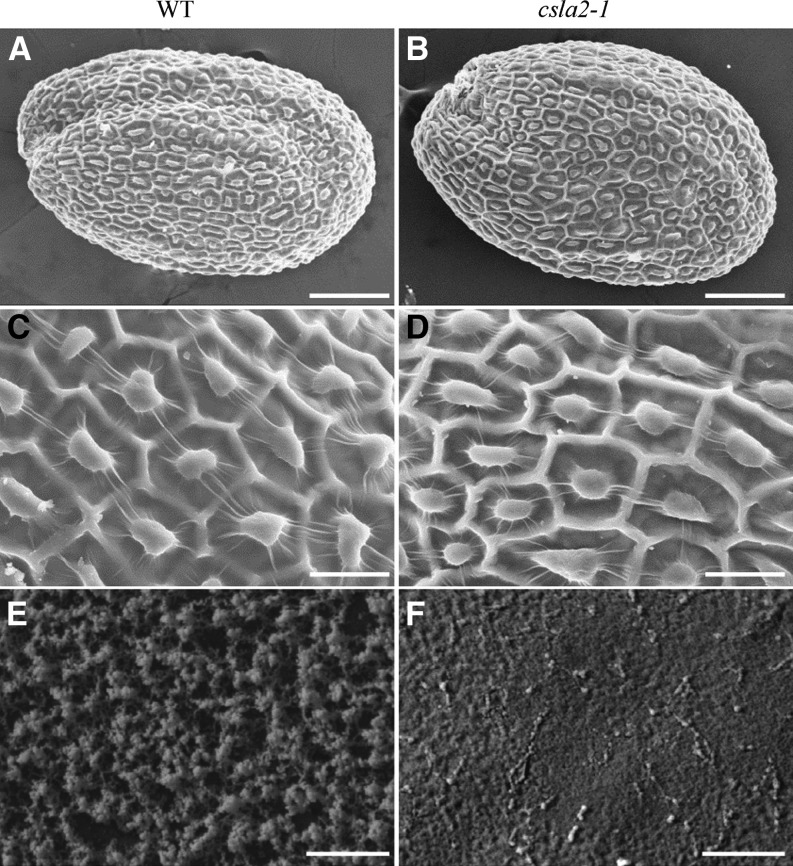
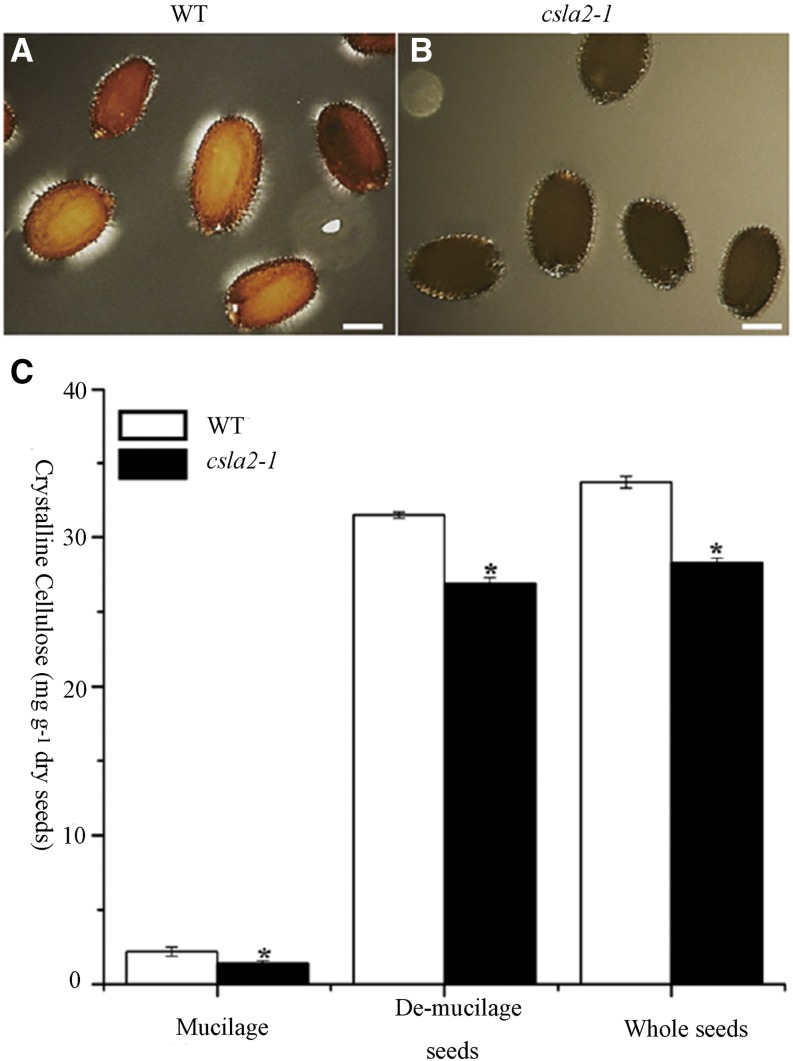
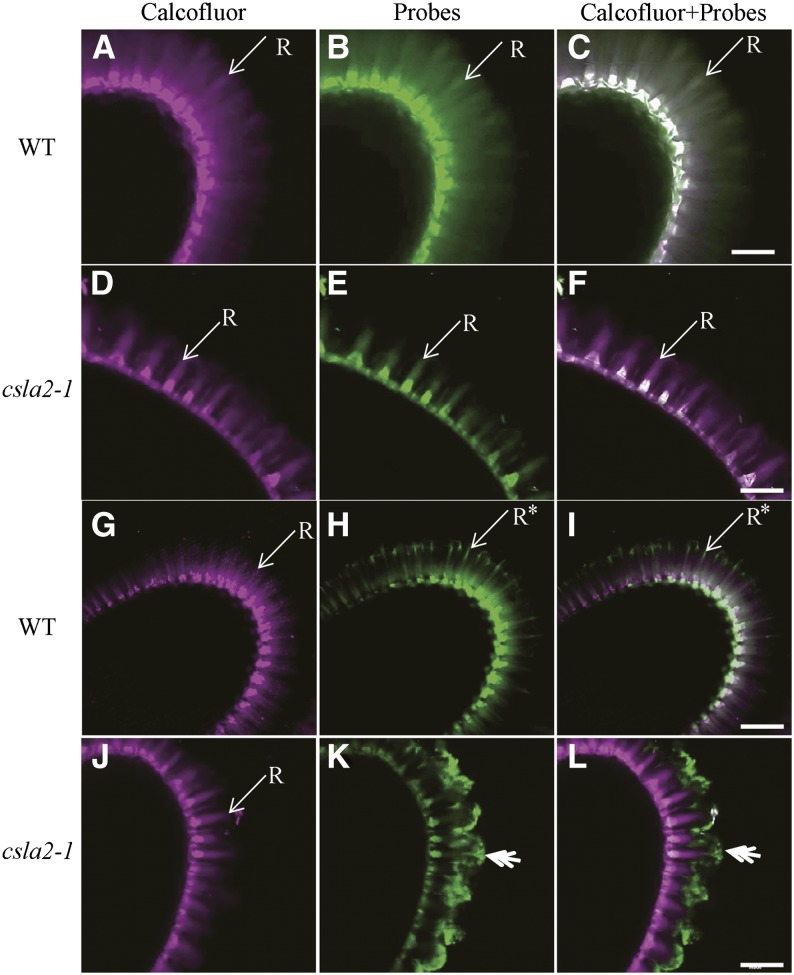
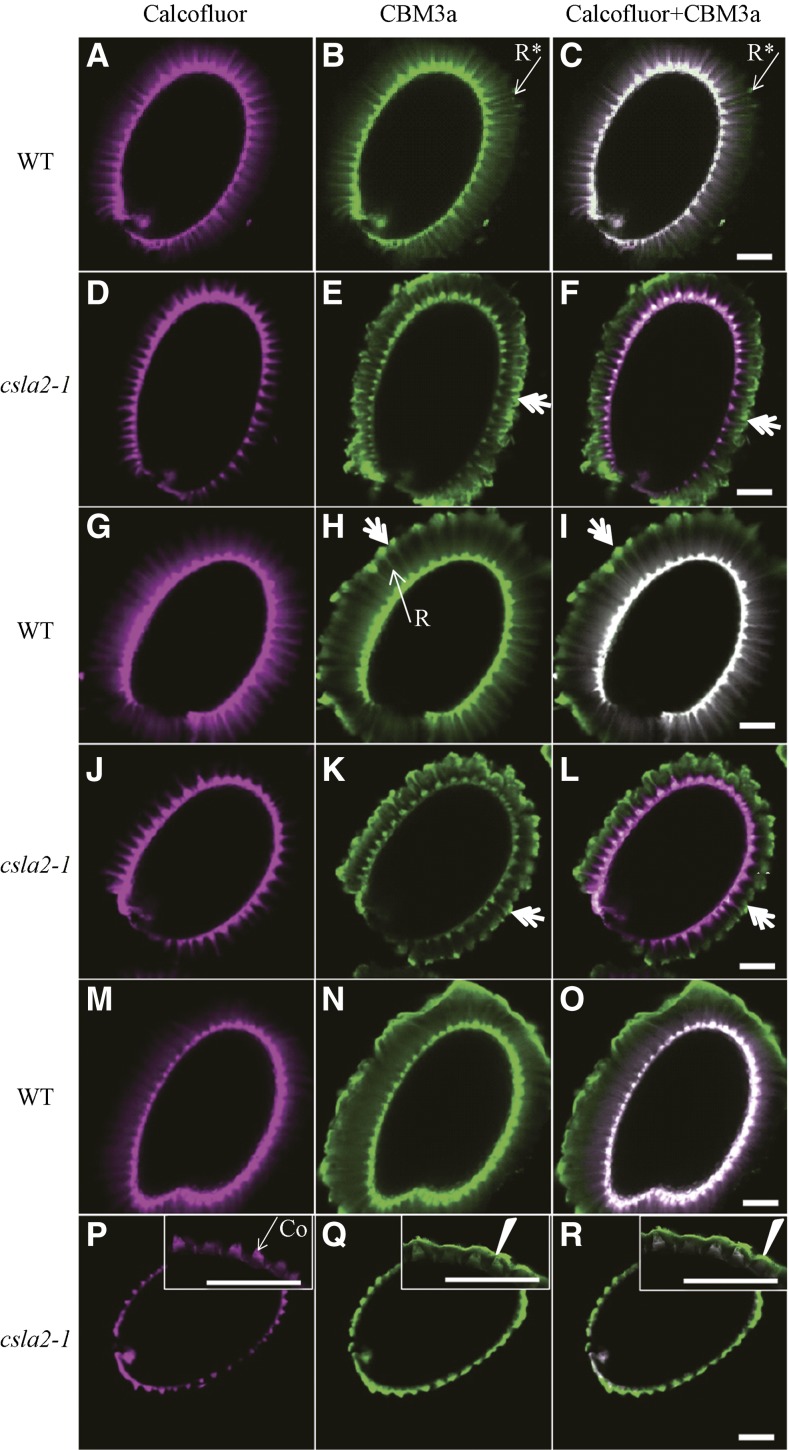
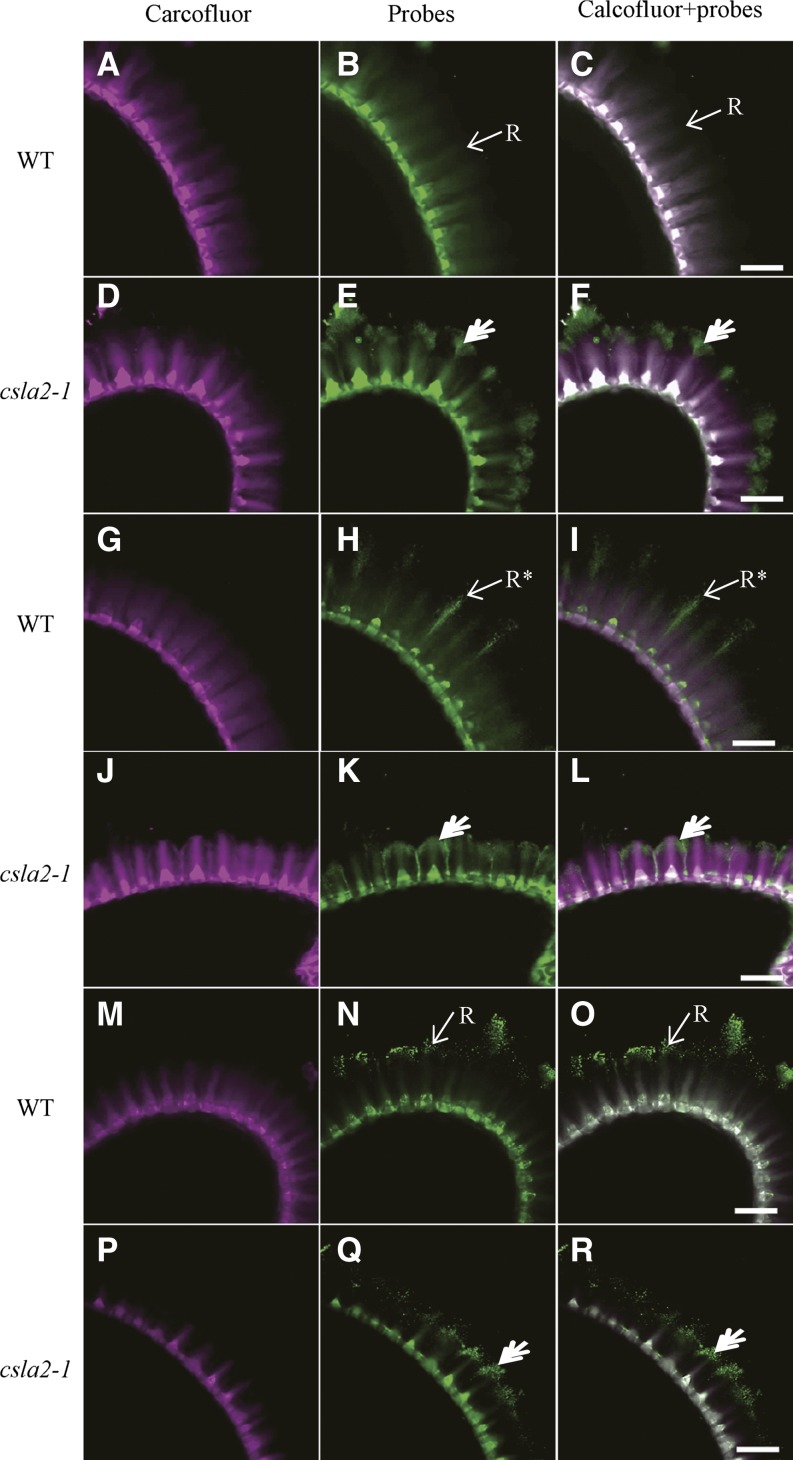
Mannans are hemicellulosic polysaccharides that are considered to have both structural and storage functions in the plant cell wall. However, it is not yet known how mannans function in Arabidopsis (Arabidopsis thaliana) seed mucilage. In this study, CELLULOSE SYNTHASE-LIKE A2 (CSLA2; At5g22740) expression was observed in several seed tissues, including the epidermal cells of developing seed coats. Disruption of CSLA2 resulted in thinner adherent mucilage halos, although the total amount of the adherent mucilage did not change compared with the wild type. This suggested that the adherent mucilage in the mutant was more compact compared with that of the wild type. In accordance with the role of CSLA2 in glucomannan synthesis, csla2-1 mucilage contained 30% less mannosyl and glucosyl content than did the wild type. No appreciable changes in the composition, structure, or macromolecular properties were observed for nonmannan polysaccharides in mutant mucilage. Biochemical analysis revealed that cellulose crystallinity was substantially reduced in csla2-1 mucilage; this was supported by the removal of most mucilage cellulose through treatment of csla2-1 seeds with endo-β-glucanase. Mutation in CSLA2 also resulted in altered spatial distribution of cellulose and an absence of birefringent cellulose microfibrils within the adherent mucilage. As with the observed changes in crystalline cellulose, the spatial distribution of pectin was also modified in csla2-1 mucilage. Taken together, our results demonstrate that glucomannans synthesized by CSLA2 are involved in modulating the structure of adherent mucilage, potentially through altering cellulose organization and crystallization.
Figures
References
-
- Beeckman T, De Rycke R, Viane R, Inzé D. (2000) Histological study of seed coat development in Arabidopsis thaliana. J Plant Res 113: 139–148
-
- Blake AW, McCartney L, Flint JE, Bolam DN, Boraston AB, Gilbert HJ, Knox JP. (2006) Understanding the biological rationale for the diversity of cellulose-directed carbohydrate-binding modules in prokaryotic enzymes. J Biol Chem 281: 29321–29329 - PubMed
-
- Blumenkrantz N, Asboe-Hansen G. (1973) New method for quantitative determination of uronic acids. Anal Biochem 54: 484–489 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
