

Cell-type-specific effects of RNase L on viral induction of beta interferon
- PMID: 24570368
- PMCID: PMC3940032
- DOI: 10.1128/mBio.00856-14
Cell-type-specific effects of RNase L on viral induction of beta interferon
Abstract
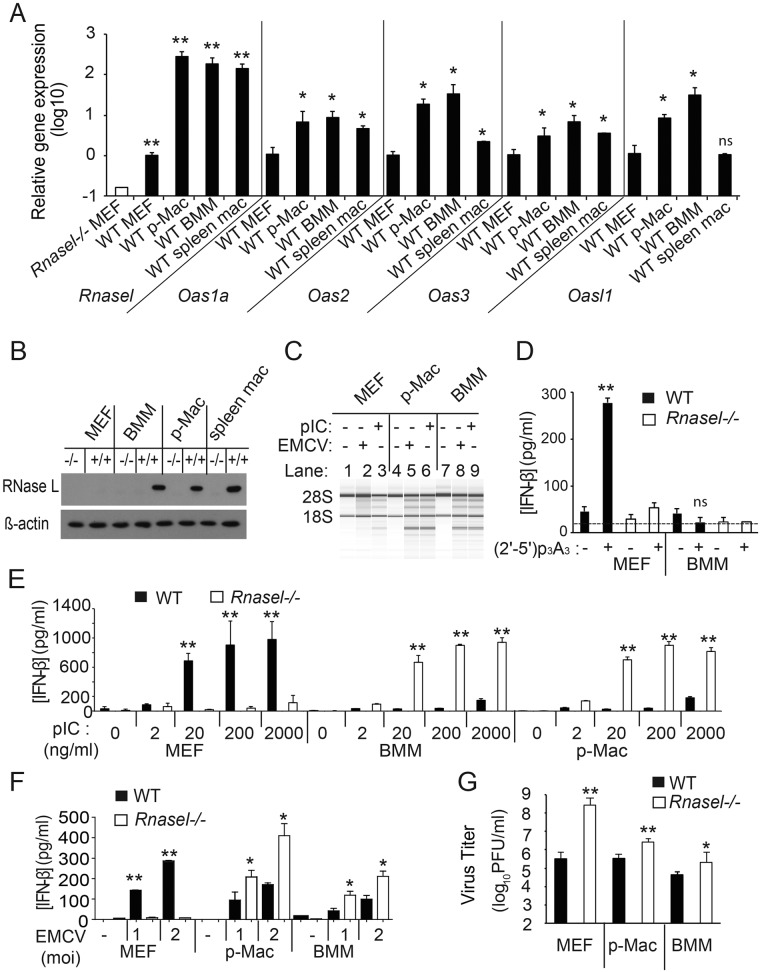
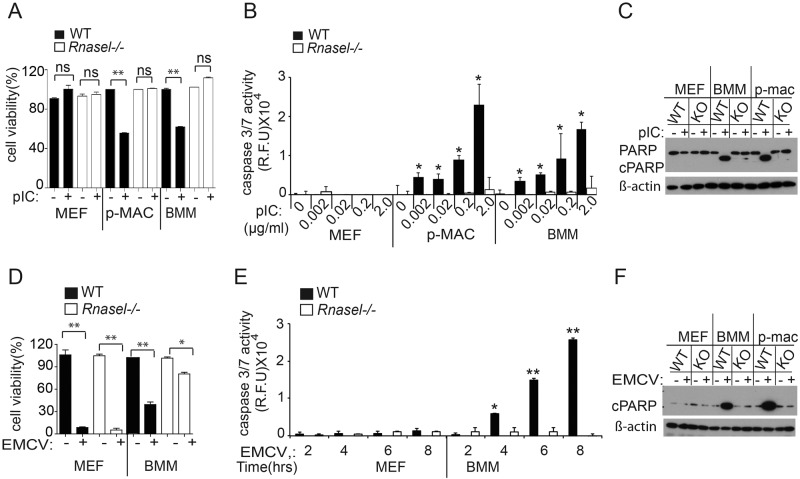
The interferon (IFN)-inducible antiviral state is mediated in part by the 2',5'-oligoadenylate (2-5A) synthetase (OAS)/RNase L system. 2-5A, produced from ATP by OAS proteins in response to viral double-stranded RNA, binds to and activates RNase L. RNase L restricts viral infections by degrading viral and cellular RNA, inducing autophagy and apoptosis, and producing RNA degradation products that amplify production of type I interferons (IFNs) through RIG-I-like receptors. However, the effects of the OAS/RNase L pathway on IFN induction in different cell types that vary in basal levels of these proteins have not been previously reported. Here we report higher basal expression of both RNase L and OAS in mouse macrophages in comparison to mouse embryonic fibroblasts (MEFs). In MEFs, RNase L gene knockout decreased induction of IFN-β by encephalomyocarditis virus infection or poly(rI):poly(rC) (pIC) transfection. In contrast, in macrophages, RNase L deletion increased (rather than decreased) induction of IFN-β by virus or pIC. RNA damage from RNase L in virus-infected macrophages is likely responsible for reducing IFN-β production. Similarly, direct activation of RNase L by transfection with 2-5A induced IFN-β in MEFs but not in macrophages. Also, viral infection or pIC transfection caused RNase L-dependent apoptosis of macrophages but not of MEFs. Our results suggest that cell-type-specific differences in basal levels of OAS and RNase L are determinants of IFN-β induction that could affect tissue protection and survival during viral infections. IMPORTANCE Type I interferons (IFNs) such as IFN-β are essential antiviral cytokines that are often required for animal survival following infections by highly pathogenic viruses. Therefore, host factors that regulate type I IFN production are critically important for animal and human health. Previously we reported that the OAS/RNase L pathway amplifies antiviral innate immunity by enhancing IFN-β production in mouse embryonic fibroblasts and in virus-infected mice. Here we report that high basal levels of OAS/RNase L in macrophages reduce, rather than increase, virus induction of IFN-β. RNA damage and apoptosis caused by RNase L were the likely reasons for the decreased IFN-β production in virus-infected macrophages. Our studies suggest that during viral infections, the OAS/RNase L pathway can either enhance or suppress IFN production, depending on the cell type. IFN regulation by RNase L is suggested to contribute to tissue protection and survival during viral infections.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
