

GIP/MZT1 proteins orchestrate nuclear shaping
- PMID: 24570680
- PMCID: PMC3916773
- DOI: 10.3389/fpls.2014.00029
GIP/MZT1 proteins orchestrate nuclear shaping
Abstract
The functional organization of the nuclear envelope (NE) is only just emerging in plants with the recent characterization of NE protein complexes and their molecular links to the actin cytoskeleton. The NE also plays a role in microtubule nucleation by recruiting γ-Tubulin Complexes (γ-TuCs) which contribute to the establishment of a robust mitotic spindle. γ-tubulin Complex Protein 3 (GCP3)-interacting proteins (GIPs) have been identified recently as integral components of γ-TuCs. GIPs have been conserved throughout evolution and are also named MZT1 (mitotic-spindle organizing protein 1). This review focuses on recent data investigating the role of GIP/MZT1 at the NE, including insights from the study of GIP partners. It also uncovers new functions for GIP/MZT1 during interphase and highlights a current view of NE-associated components which are critical for nuclear shaping during both cell division and differentiation.
Keywords: Arabidopsis thaliana; gamma-tubulin complex; nuclear envelope proteins; nucleocytoplasmic continuum; spindle assembly.
Figures
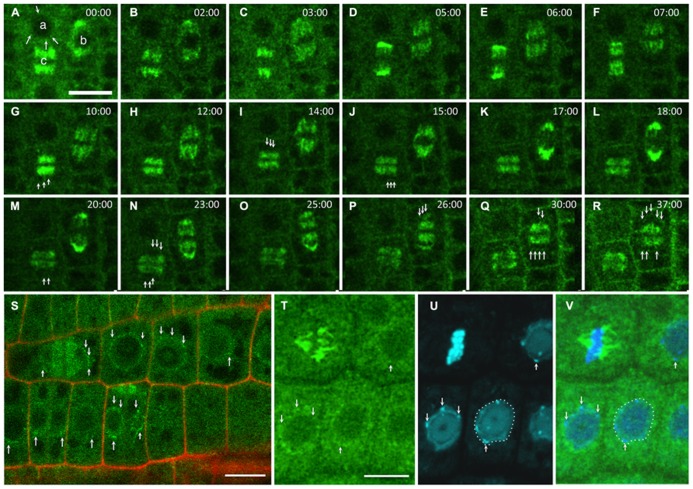
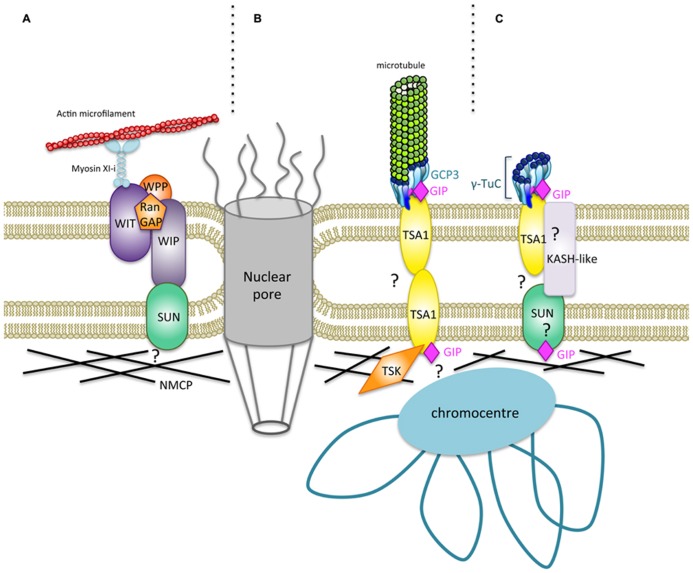
References
-
- Canaday J., Brochot A. L., Seltzer V., Herzog E., Evrard J. L., Schmit A. C. (2005) “Microtubule assembly in higher plants,” in Recent Research Developments in Molecular Biology Vol. 2 ed. Pandalai S. G. (Trivandrum, India: Research Signpost) 103–119
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials