

The hippocampal CA2 region is essential for social memory
- PMID: 24572357
- PMCID: PMC4000264
- DOI: 10.1038/nature13028
The hippocampal CA2 region is essential for social memory
Abstract
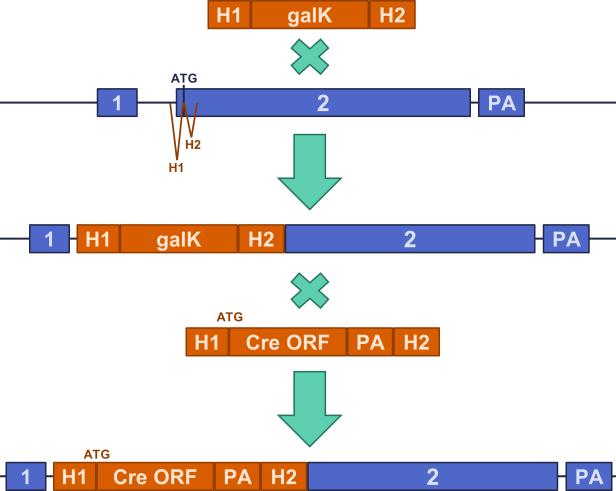
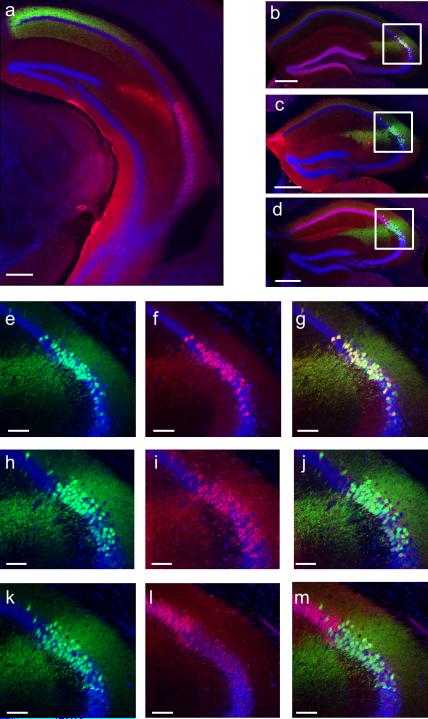
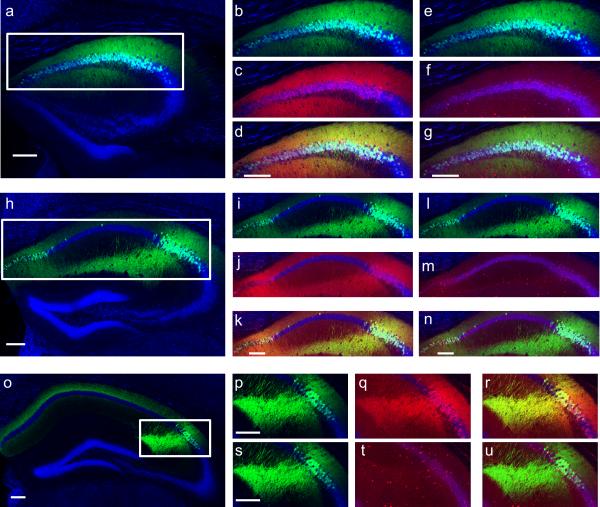
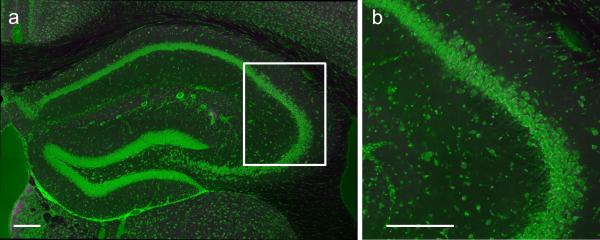
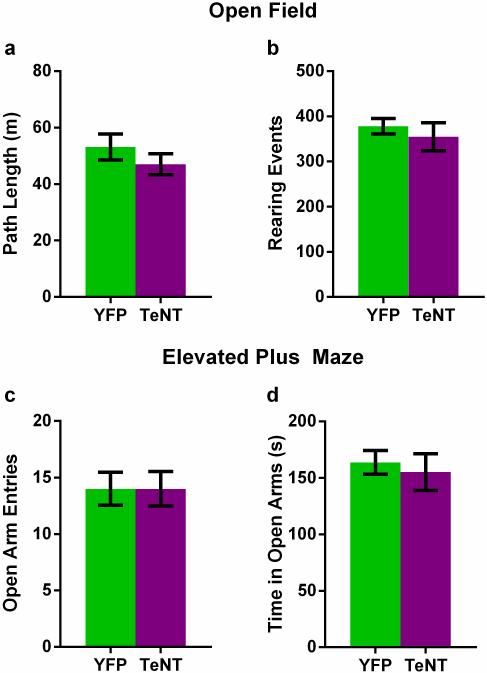
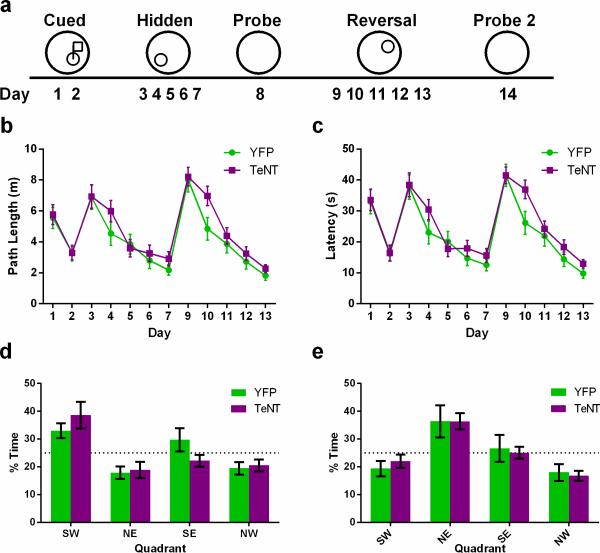
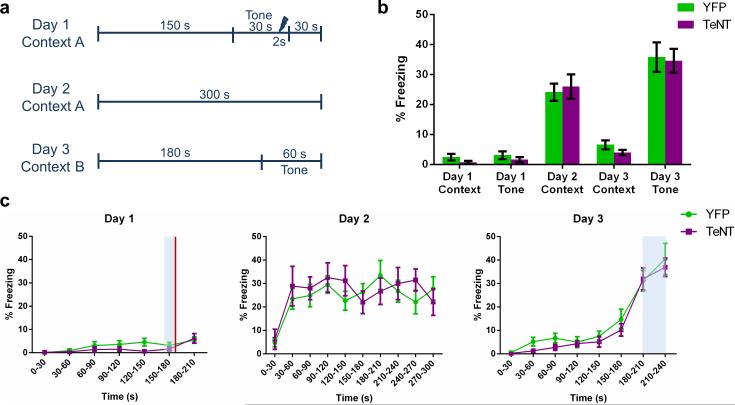
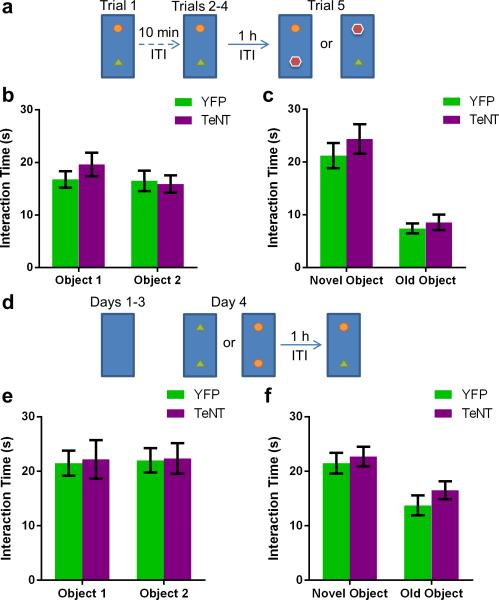
The hippocampus is critical for encoding declarative memory, our repository of knowledge of who, what, where and when. Mnemonic information is processed in the hippocampus through several parallel routes involving distinct subregions. In the classic trisynaptic pathway, information proceeds from entorhinal cortex (EC) to dentate gyrus to CA3 and then to CA1, the main hippocampal output. Genetic lesions of EC (ref. 3) and hippocampal dentate gyrus (ref. 4), CA3 (ref. 5) and CA1 (ref. 6) regions have revealed their distinct functions in learning and memory. In contrast, little is known about the role of CA2, a relatively small area interposed between CA3 and CA1 that forms the nexus of a powerful disynaptic circuit linking EC input with CA1 output. Here we report a novel transgenic mouse line that enabled us to selectively examine the synaptic connections and behavioural role of the CA2 region in adult mice. Genetically targeted inactivation of CA2 pyramidal neurons caused a pronounced loss of social memory--the ability of an animal to remember a conspecific--with no change in sociability or several other hippocampus-dependent behaviours, including spatial and contextual memory. These behavioural and anatomical results thus reveal CA2 as a critical hub of sociocognitive memory processing.
Figures
Similar articles
-
Strong CA2 pyramidal neuron synapses define a powerful disynaptic cortico-hippocampal loop.Neuron. 2010 May 27;66(4):560-72. doi: 10.1016/j.neuron.2010.04.013. Neuron. 2010. PMID: 20510860 Free PMC article.
-
RGS14 is a natural suppressor of both synaptic plasticity in CA2 neurons and hippocampal-based learning and memory.Proc Natl Acad Sci U S A. 2010 Sep 28;107(39):16994-8. doi: 10.1073/pnas.1005362107. Epub 2010 Sep 13. Proc Natl Acad Sci U S A. 2010. PMID: 20837545 Free PMC article.
-
Hippocampal area CA2: an emerging modulatory gateway in the hippocampal circuit.Exp Brain Res. 2018 Apr;236(4):919-931. doi: 10.1007/s00221-018-5187-5. Epub 2018 Jan 31. Exp Brain Res. 2018. PMID: 29387909 Review.
-
Conditional Deletion of Hippocampal CA2/CA3a Oxytocin Receptors Impairs the Persistence of Long-Term Social Recognition Memory in Mice.J Neurosci. 2018 Jan 31;38(5):1218-1231. doi: 10.1523/JNEUROSCI.1896-17.2017. Epub 2017 Dec 26. J Neurosci. 2018. PMID: 29279308 Free PMC article.
-
CA2: A Highly Connected Intrahippocampal Relay.Annu Rev Neurosci. 2020 Jul 8;43:55-72. doi: 10.1146/annurev-neuro-080719-100343. Epub 2019 Dec 24. Annu Rev Neurosci. 2020. PMID: 31874067 Review.
Cited by
-
MMP9 modulation improves specific neurobehavioral deficits in a mouse model of Alzheimer's disease.BMC Neurosci. 2021 May 25;22(1):39. doi: 10.1186/s12868-021-00643-2. BMC Neurosci. 2021. PMID: 34034683 Free PMC article.
-
Autistic-like traits in laboratory rodents exposed to phthalic acid esters during early development - an animal model of autism?Physiol Res. 2021 Jul 12;70(3):345-361. doi: 10.33549/physiolres.934570. Epub 2021 May 12. Physiol Res. 2021. PMID: 33982578 Free PMC article. Review.
-
Oxytocin Ameliorates Impaired Behaviors of High Fat Diet-Induced Obese Mice.Front Endocrinol (Lausanne). 2020 Jul 3;11:379. doi: 10.3389/fendo.2020.00379. eCollection 2020. Front Endocrinol (Lausanne). 2020. PMID: 32719656 Free PMC article.
-
Hippocampal interlamellar cell-cell connectome that counts.J Cell Physiol. 2022 Nov;237(11):4037-4048. doi: 10.1002/jcp.30868. Epub 2022 Sep 5. J Cell Physiol. 2022. PMID: 36063532 Free PMC article. Review.
-
Disruption of the autism gene and chromatin regulator KDM5A alters hippocampal cell identity.Sci Adv. 2023 Nov 24;9(47):eadi0074. doi: 10.1126/sciadv.adi0074. Epub 2023 Nov 22. Sci Adv. 2023. PMID: 37992166 Free PMC article.
References
-
- van Strien NM, Cappaert NL, Witter MP. The anatomy of memory: an interactive overview of the parahippocampal-hippocampal network. Nat. Rev. Neurosci. 2009;10:272–282. - PubMed
-
- Suh J, Rivest AJ, Nakashiba T, Tominaga T, Tonegawa S. Entorhinal cortex layer III input to the hippocampus is crucial for temporal association memory. Science. 2011;334:1415–20. - PubMed
-
- Nakashiba T, Young JZ, McHugh TJ, Buhl DL, Tonegawa S. Transgenic inhibition of synaptic transmission reveals role of CA3 output in hippocampal learning. Science. 2008;319:1260–4. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
