

A stem-loop structure directs oskar mRNA to microtubule minus ends
- PMID: 24572808
- PMCID: PMC3964905
- DOI: 10.1261/rna.041566.113
A stem-loop structure directs oskar mRNA to microtubule minus ends
Abstract
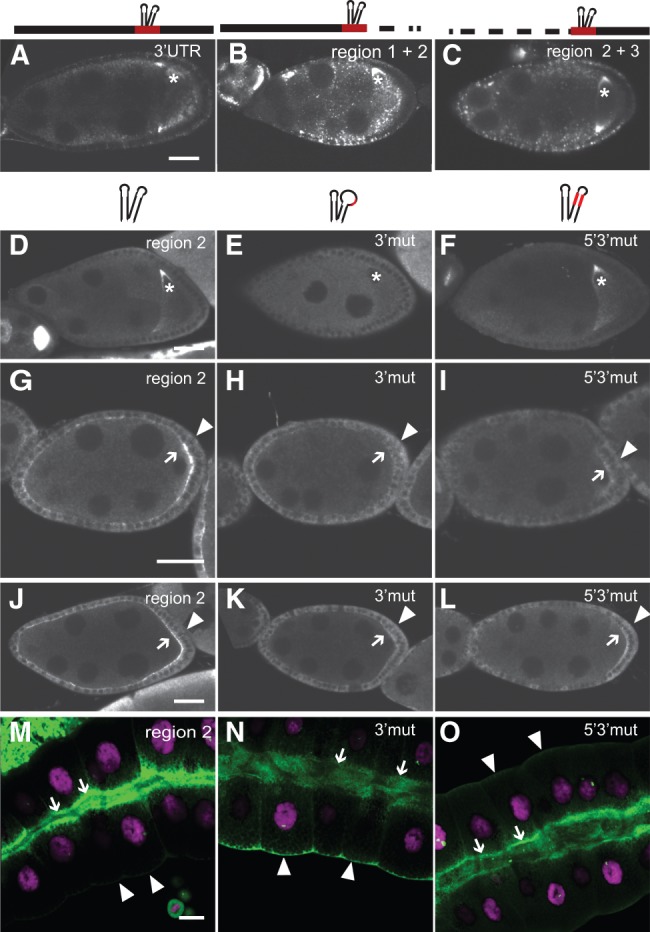
mRNA transport coupled with translational control underlies the intracellular localization of many proteins in eukaryotic cells. This is exemplified in Drosophila, where oskar mRNA transport and translation at the posterior pole of the oocyte direct posterior patterning of the embryo. oskar localization is a multistep process. Within the oocyte, a spliced oskar localization element (SOLE) targets oskar mRNA for plus end-directed transport by kinesin-1 to the posterior pole. However, the signals mediating the initial minus end-directed, dynein-dependent transport of the mRNA from nurse cells into the oocyte have remained unknown. Here, we show that a 67-nt stem-loop in the oskar 3' UTR promotes oskar mRNA delivery to the developing oocyte and that it shares functional features with the fs(1)K10 oocyte localization signal. Thus, two independent cis-acting signals, the oocyte entry signal (OES) and the SOLE, mediate sequential dynein- and kinesin-dependent phases of oskar mRNA transport during oogenesis. The OES also promotes apical localization of injected RNAs in blastoderm stage embryos, another dynein-mediated process. Similarly, when ectopically expressed in polarized cells of the follicular epithelium or salivary glands, reporter RNAs bearing the oskar OES are apically enriched, demonstrating that this element promotes mRNA localization independently of cell type. Our work sheds new light on how oskar mRNA is trafficked during oogenesis and the RNA features that mediate minus end-directed transport.
Keywords: Drosophila; mRNA localization; oocyte; oogenesis; osk; oskar.
Figures



Similar articles
-
Different roles for the adjoining and structurally similar A-rich and poly(A) domains of oskar mRNA: Only the A-rich domain is required for oskar noncoding RNA function, which includes MTOC positioning.Dev Biol. 2021 Aug;476:117-127. doi: 10.1016/j.ydbio.2021.03.021. Epub 2021 Mar 31. Dev Biol. 2021. PMID: 33798537 Free PMC article.
-
Multiple cis-acting signals, some weak by necessity, collectively direct robust transport of oskar mRNA to the oocyte.J Cell Sci. 2017 Sep 15;130(18):3060-3071. doi: 10.1242/jcs.202069. Epub 2017 Jul 31. J Cell Sci. 2017. PMID: 28760927 Free PMC article.
-
Polar transport in the Drosophila oocyte requires Dynein and Kinesin I cooperation.Curr Biol. 2002 Dec 10;12(23):1971-81. doi: 10.1016/s0960-9822(02)01302-7. Curr Biol. 2002. PMID: 12477385
-
Microtubule-based motor-mediated mRNA localization in Drosophila oocytes and embryos.Biochem Soc Trans. 2011 Oct;39(5):1197-201. doi: 10.1042/BST0391197. Biochem Soc Trans. 2011. PMID: 21936788 Review.
-
Translational regulation and RNA localization in Drosophila oocytes and embryos.Annu Rev Genet. 2001;35:365-406. doi: 10.1146/annurev.genet.35.102401.090756. Annu Rev Genet. 2001. PMID: 11700288 Review.
Cited by
-
Hepatitis B virus genotypes and genome characteristics in China.World J Gastroenterol. 2015 Jun 7;21(21):6684-97. doi: 10.3748/wjg.v21.i21.6684. World J Gastroenterol. 2015. PMID: 26074707 Free PMC article.
-
Liquid-to-solid phase transition of oskar ribonucleoprotein granules is essential for their function in Drosophila embryonic development.Cell. 2022 Apr 14;185(8):1308-1324.e23. doi: 10.1016/j.cell.2022.02.022. Epub 2022 Mar 23. Cell. 2022. PMID: 35325593 Free PMC article.
-
Localization of oskar mRNA by agglomeration in ribonucleoprotein granules.PLoS Genet. 2023 Aug 25;19(8):e1010877. doi: 10.1371/journal.pgen.1010877. eCollection 2023 Aug. PLoS Genet. 2023. PMID: 37624861 Free PMC article.
-
An RNA-binding atypical tropomyosin recruits kinesin-1 dynamically to oskar mRNPs.EMBO J. 2017 Feb 1;36(3):319-333. doi: 10.15252/embj.201696038. Epub 2016 Dec 27. EMBO J. 2017. PMID: 28028052 Free PMC article.
-
Opposing roles for Egalitarian and Staufen in transport, anchoring and localization of oskar mRNA in the Drosophila oocyte.PLoS Genet. 2021 Apr 2;17(4):e1009500. doi: 10.1371/journal.pgen.1009500. eCollection 2021 Apr. PLoS Genet. 2021. PMID: 33798193 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials