

ASIC1a activation enhances inhibition in the basolateral amygdala and reduces anxiety
- PMID: 24573273
- PMCID: PMC3935079
- DOI: 10.1523/JNEUROSCI.4009-13.2014
ASIC1a activation enhances inhibition in the basolateral amygdala and reduces anxiety
Abstract
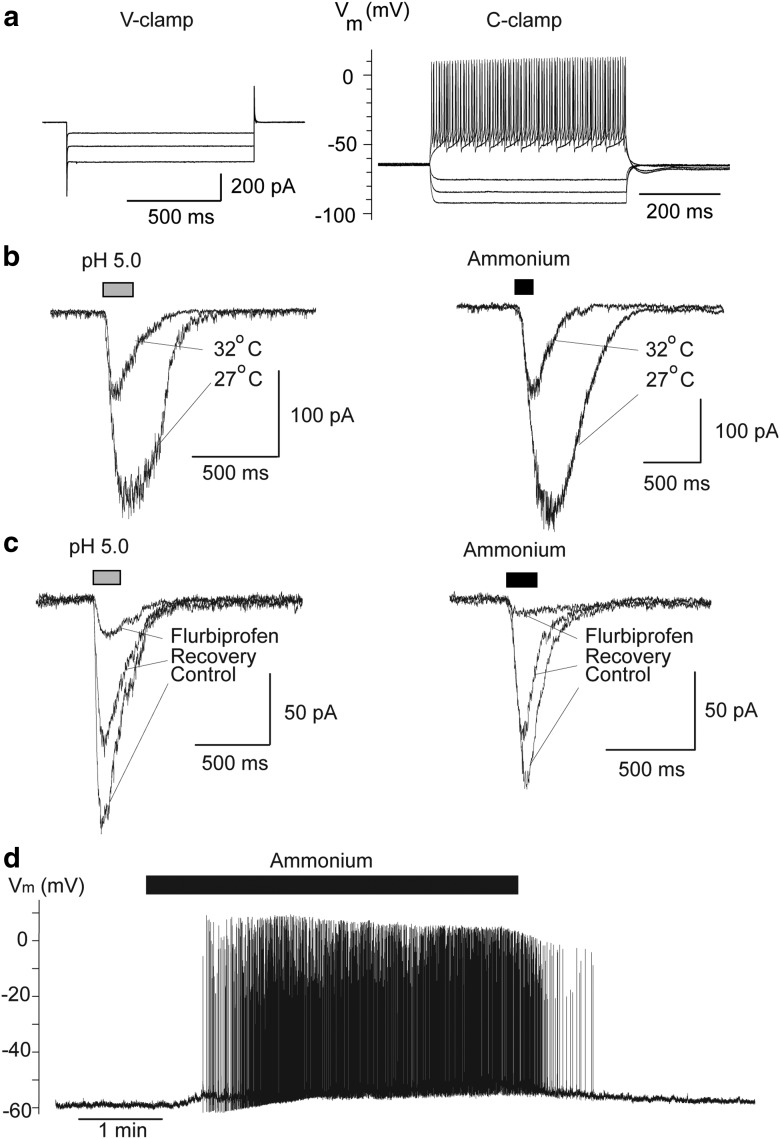
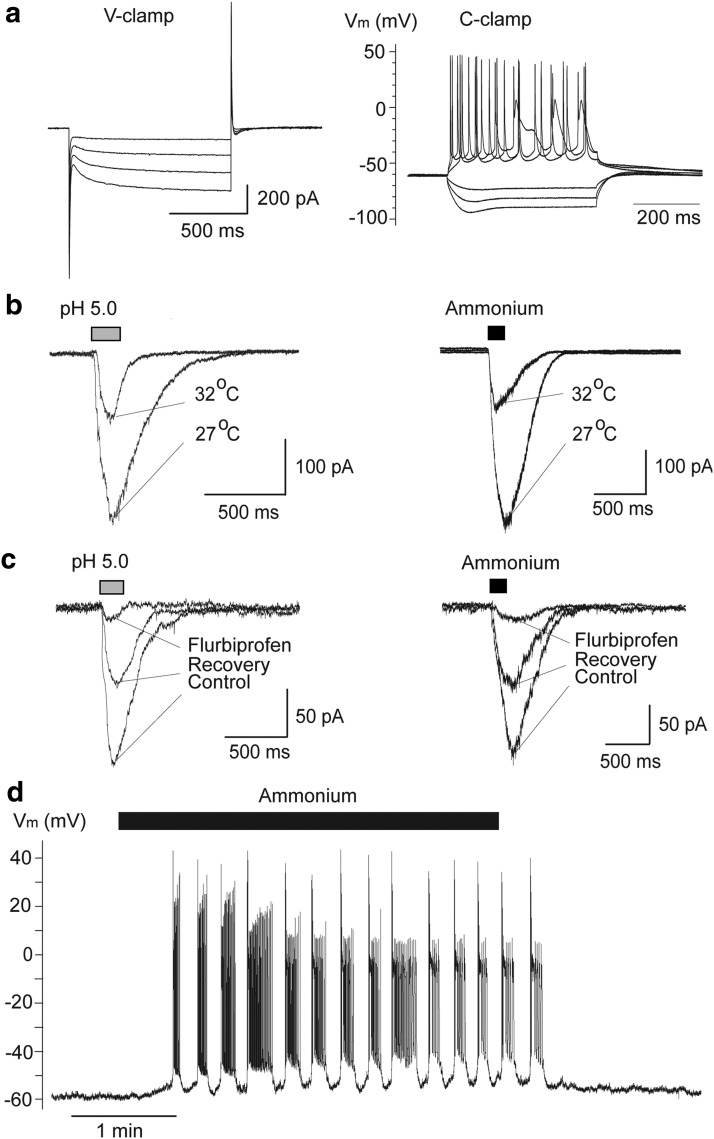
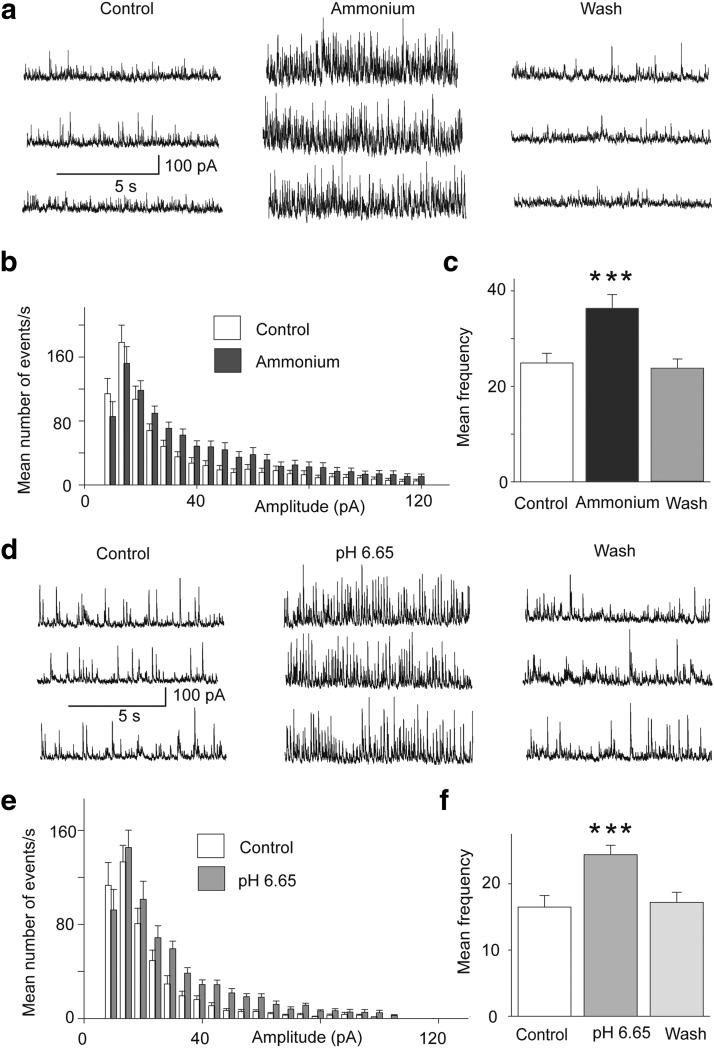
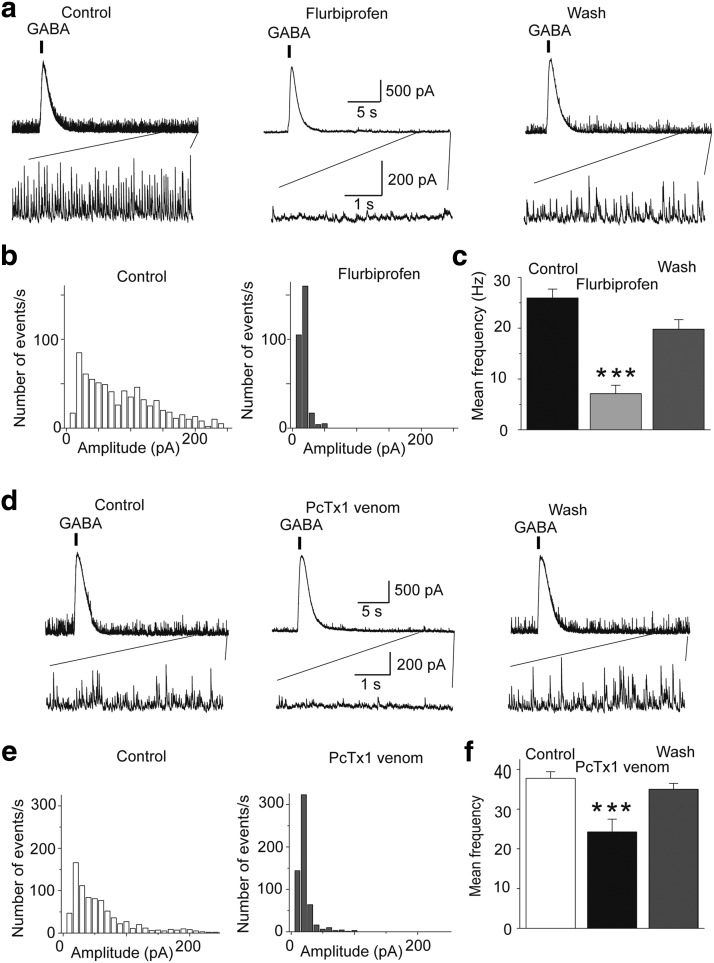
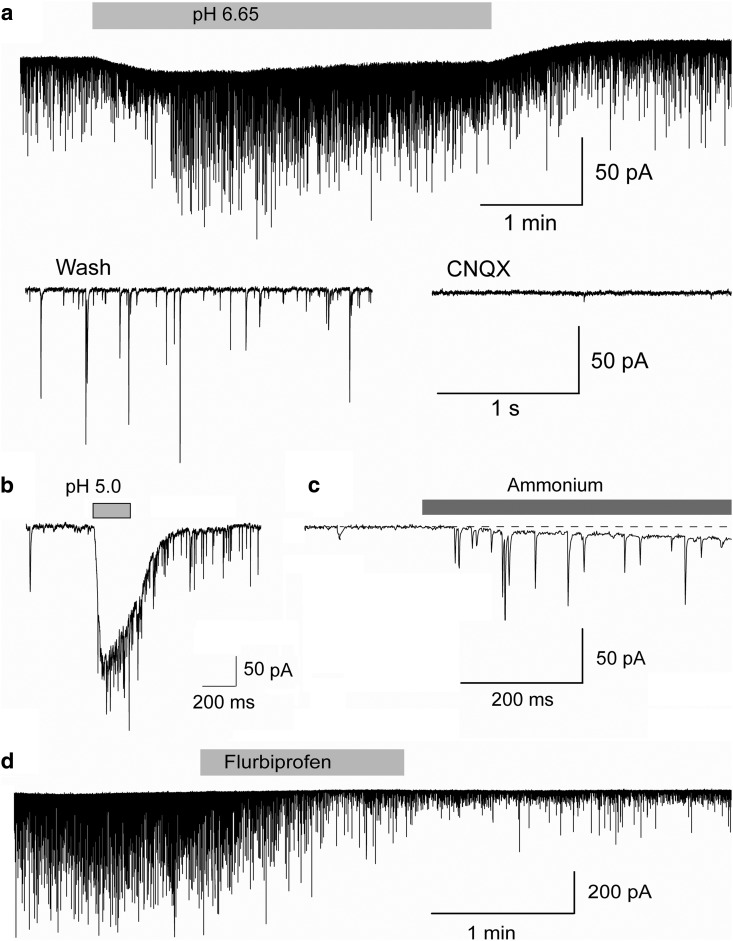
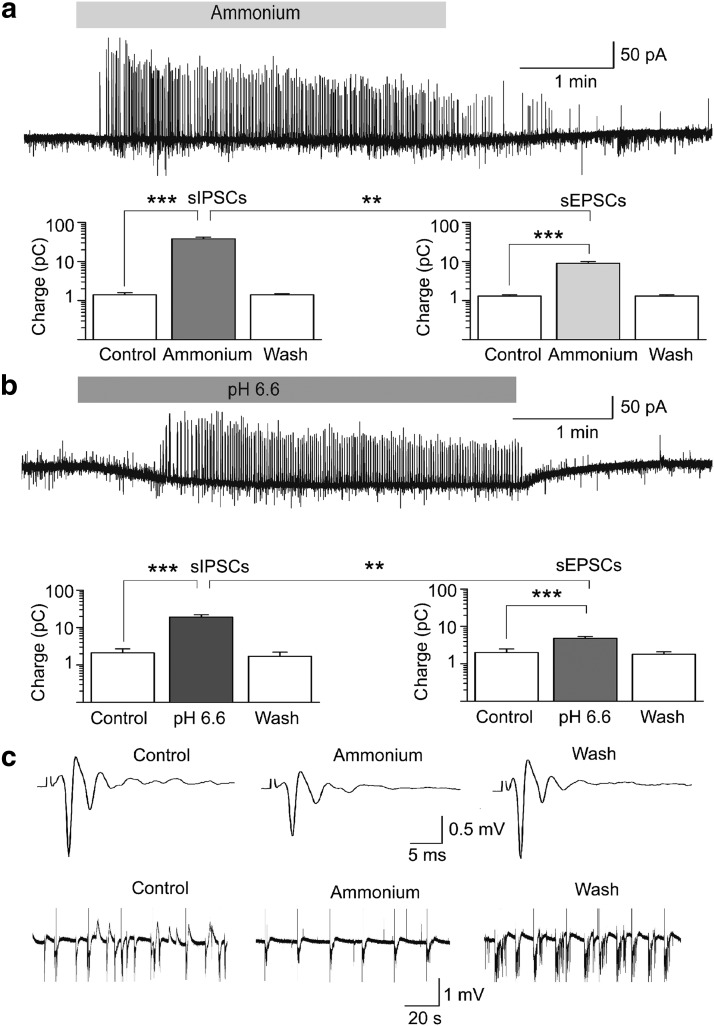
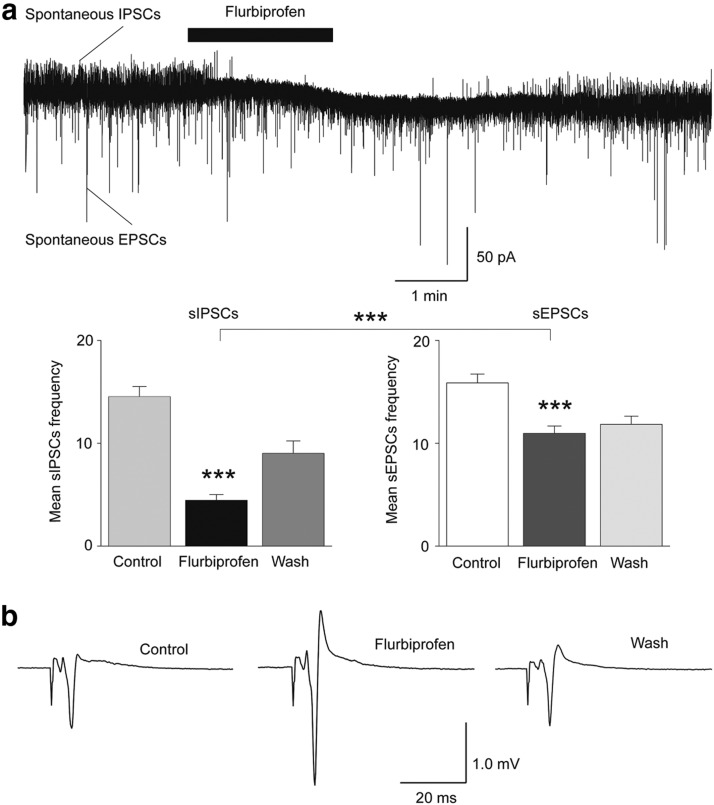
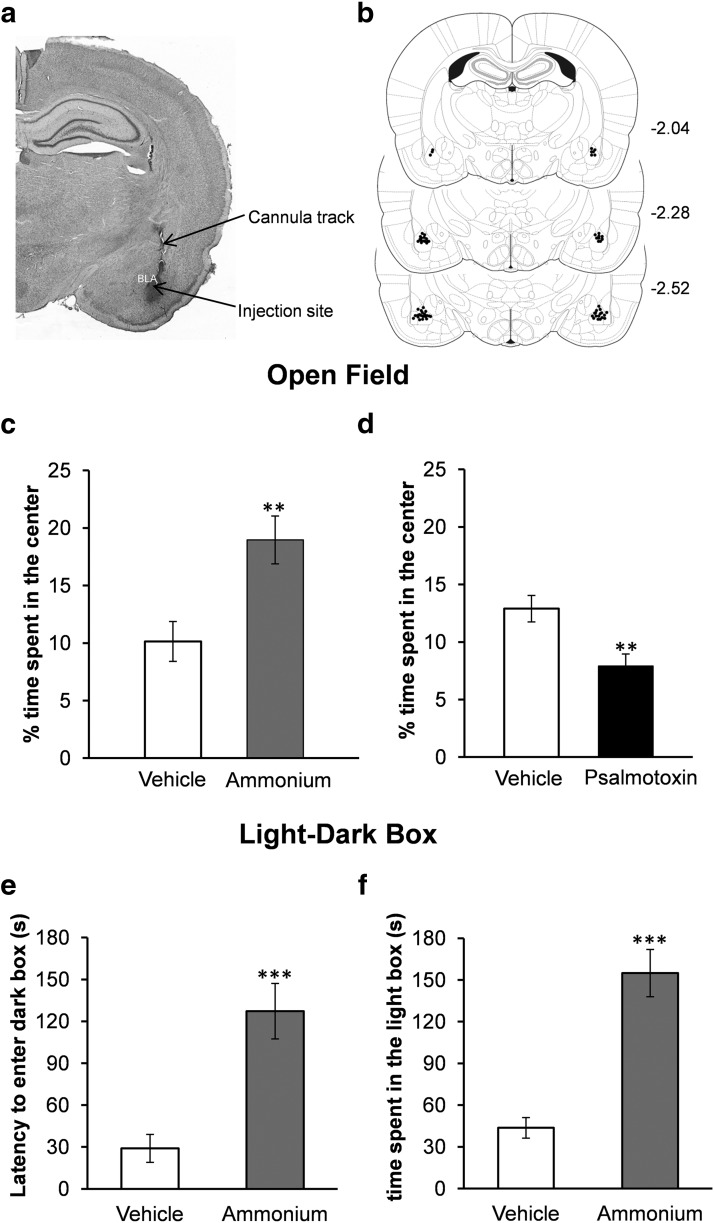
The discovery that even small changes in extracellular acidity can alter the excitability of neuronal networks via activation of acid-sensing ion channels (ASICs) could have therapeutic application in a host of neurological and psychiatric illnesses. Recent evidence suggests that activation of ASIC1a, a subtype of ASICs that is widely distributed in the brain, is necessary for the expression of fear and anxiety. Antagonists of ASIC1a, therefore, have been proposed as a potential treatment for anxiety. The basolateral amygdala (BLA) is central to fear generation, and anxiety disorders are characterized by BLA hyperexcitability. To better understand the role of ASIC1a in anxiety, we attempted to provide a direct assessment of whether ASIC1a activation increases BLA excitability. In rat BLA slices, activation of ASIC1a by low pH or ammonium elicited inward currents in both interneurons and principal neurons, and increased spontaneous IPSCs recorded from principal cells significantly more than spontaneous EPSCs. Epileptiform activity induced by high potassium and low magnesium was suppressed by ammonium. Antagonism of ASIC1a decreased spontaneous IPSCs more than EPSCs, and increased the excitability of the BLA network, as reflected by the pronounced increase of evoked field potentials, suggesting that ASIC1a channels are active in the basal state. In vivo activation or blockade of ASIC1a in the BLA suppressed or increased, respectively, anxiety-like behavior. Thus, in the rat BLA, ASIC1a has an inhibitory and anxiolytic function. The discovery of positive ASIC1a modulators may hold promise for the treatment of anxiety disorders.
Keywords: ASIC1a; GABAergic inhibition; acid-sensing ion channels; anxiety; basolateral amygdala; epileptiform activity.
Figures
Similar articles
-
Lateral paracapsular GABAergic synapses in the basolateral amygdala contribute to the anxiolytic effects of beta 3 adrenoceptor activation.Neuropsychopharmacology. 2010 Aug;35(9):1886-96. doi: 10.1038/npp.2010.59. Epub 2010 Apr 21. Neuropsychopharmacology. 2010. PMID: 20410872 Free PMC article.
-
Increased inhibitory activity in the basolateral amygdala and decreased anxiety during estrus: A potential role for ASIC1a channels.Brain Res. 2021 Nov 1;1770:147628. doi: 10.1016/j.brainres.2021.147628. Epub 2021 Aug 26. Brain Res. 2021. PMID: 34454948
-
Presynaptic facilitation of glutamate release in the basolateral amygdala: a mechanism for the anxiogenic and seizurogenic function of GluK1 receptors.Neuroscience. 2012 Sep 27;221:157-69. doi: 10.1016/j.neuroscience.2012.07.006. Epub 2012 Jul 13. Neuroscience. 2012. PMID: 22796081 Free PMC article.
-
The basolateral amygdala γ-aminobutyric acidergic system in health and disease.J Neurosci Res. 2016 Jun;94(6):548-67. doi: 10.1002/jnr.23690. Epub 2015 Nov 19. J Neurosci Res. 2016. PMID: 26586374 Free PMC article. Review.
-
Mechanisms regulating GABAergic inhibitory transmission in the basolateral amygdala: implications for epilepsy and anxiety disorders.Amino Acids. 2007;32(3):305-15. doi: 10.1007/s00726-006-0415-x. Epub 2006 Oct 18. Amino Acids. 2007. PMID: 17048126 Review.
Cited by
-
Recent advances in anxiety disorders: Focus on animal models and pathological mechanisms.Animal Model Exp Med. 2023 Dec;6(6):559-572. doi: 10.1002/ame2.12360. Epub 2023 Nov 28. Animal Model Exp Med. 2023. PMID: 38013621 Free PMC article. Review.
-
Postsynaptic Targeting and Mobility of Membrane Surface-Localized hASIC1a.Neurosci Bull. 2021 Feb;37(2):145-165. doi: 10.1007/s12264-020-00581-9. Epub 2020 Sep 30. Neurosci Bull. 2021. PMID: 32996060 Free PMC article.
-
Divalent cation and chloride ion sites of chicken acid sensing ion channel 1a elucidated by x-ray crystallography.PLoS One. 2018 Aug 29;13(8):e0202134. doi: 10.1371/journal.pone.0202134. eCollection 2018. PLoS One. 2018. PMID: 30157194 Free PMC article.
-
ASICs and neuropeptides.Neuropharmacology. 2015 Jul;94:36-41. doi: 10.1016/j.neuropharm.2014.12.012. Epub 2015 Jan 12. Neuropharmacology. 2015. PMID: 25592215 Free PMC article. Review.
-
Antiseizure and Neuroprotective Efficacy of Midazolam in Comparison with Tezampanel (LY293558) against Soman-Induced Status Epilepticus.Toxics. 2022 Jul 22;10(8):409. doi: 10.3390/toxics10080409. Toxics. 2022. PMID: 35893842 Free PMC article.
References
-
- Aroniadou-Anderjaska V, Pidoplichko VI, Figueiredo TH, Almeida-Suhett CP, Prager EM, Braga MF. Presynaptic facilitation of glutamate release in the basolateral amygdala: a mechanism for the anxiogenic and seizurogenic function of GluK1 receptors. Neuroscience. 2012;221:157–169. doi: 10.1016/j.neuroscience.2012.07.006. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical