

Proteasome modulates positive and negative translational regulators in long-term synaptic plasticity
- PMID: 24573276
- PMCID: PMC3935082
- DOI: 10.1523/JNEUROSCI.3291-13.2014
Proteasome modulates positive and negative translational regulators in long-term synaptic plasticity
Abstract
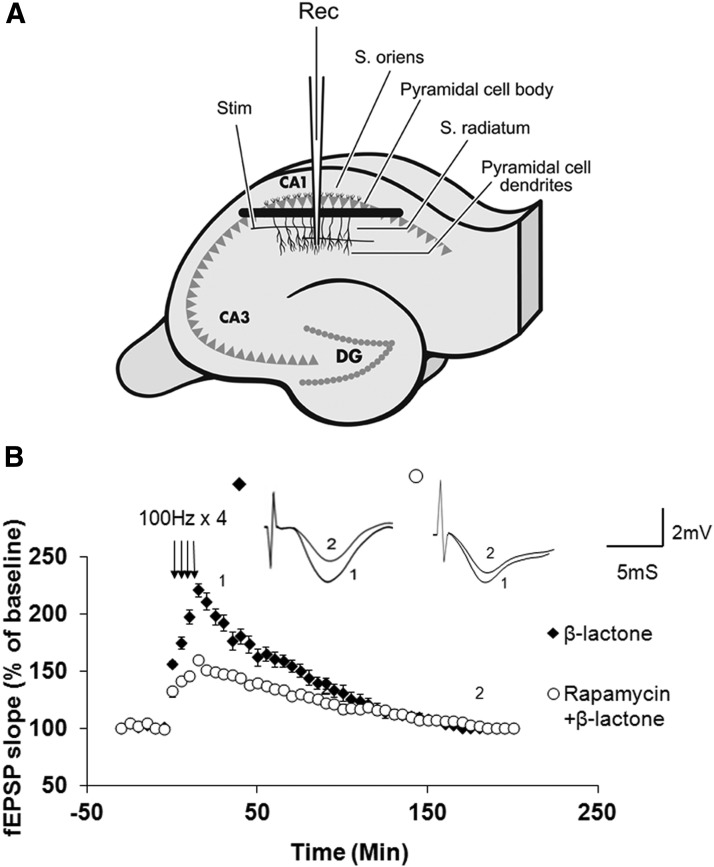
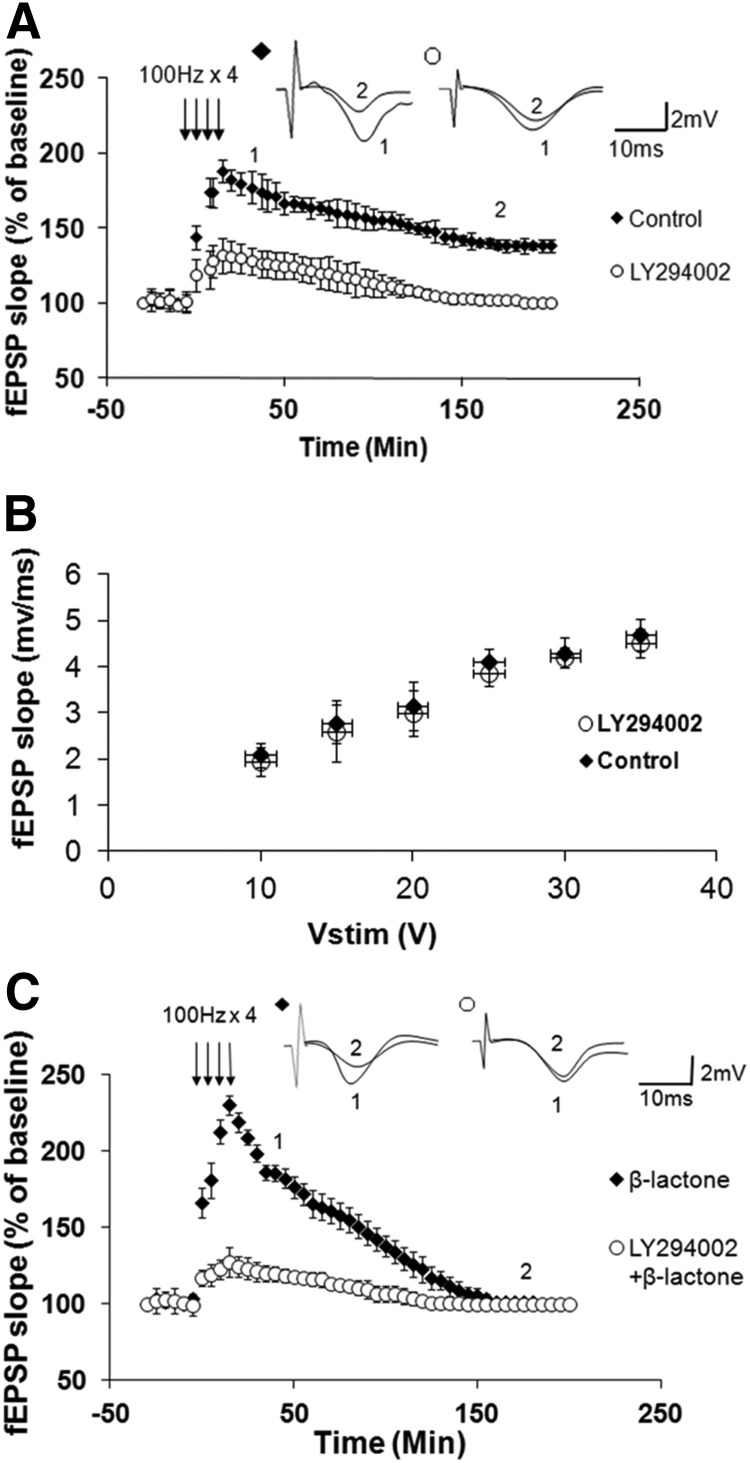
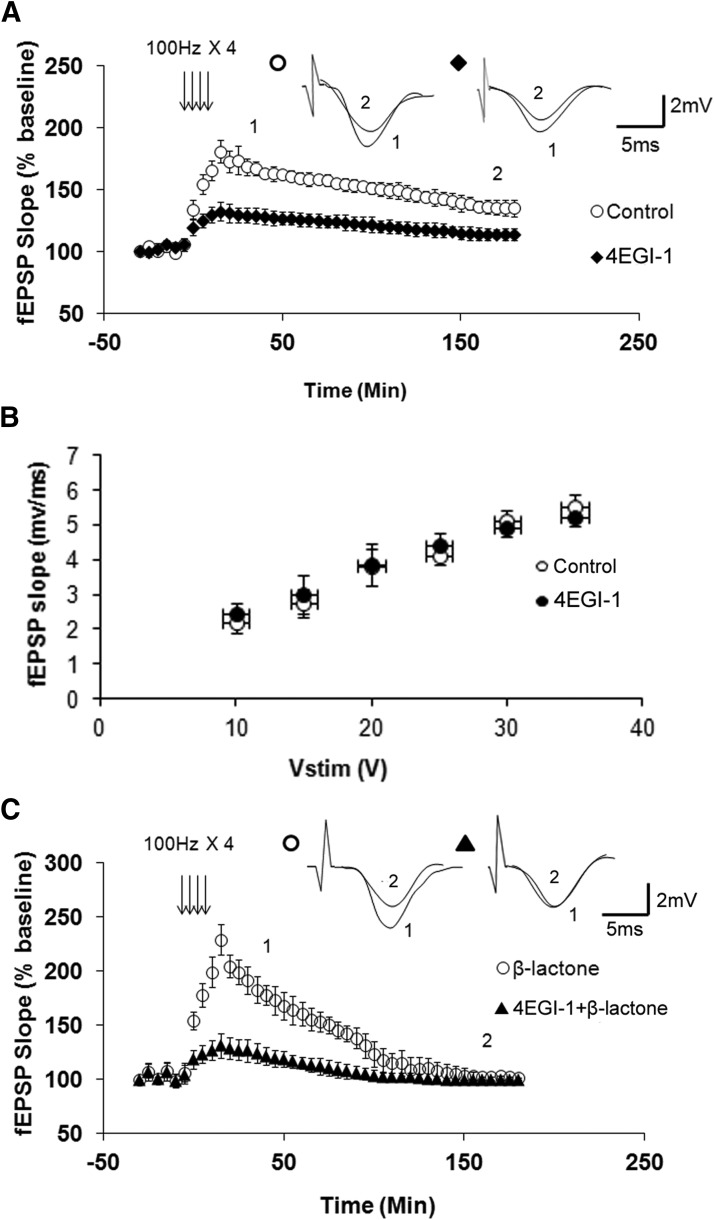
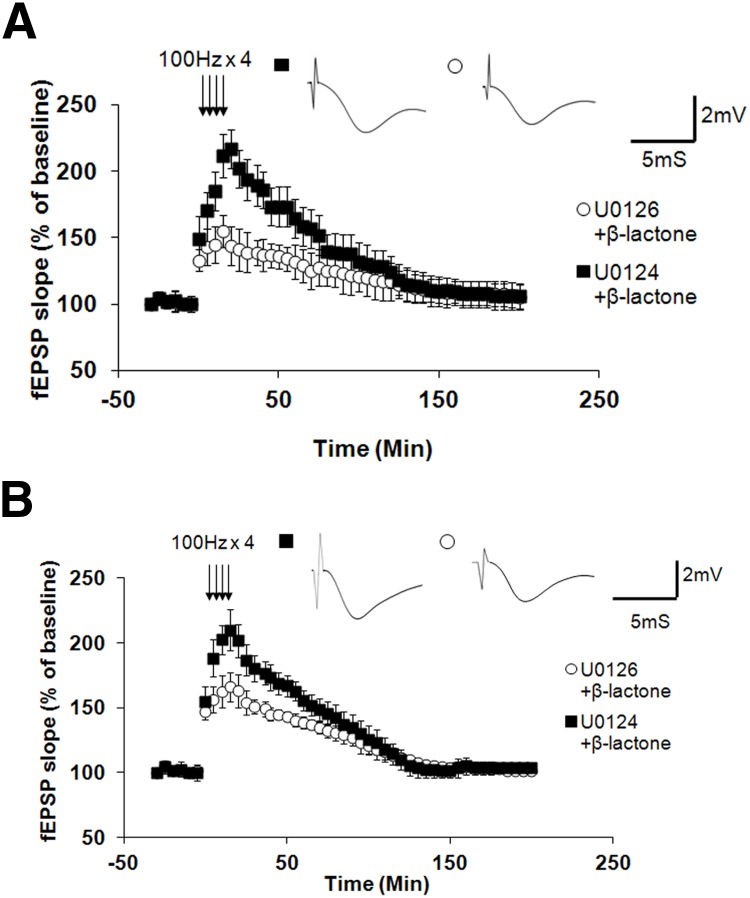
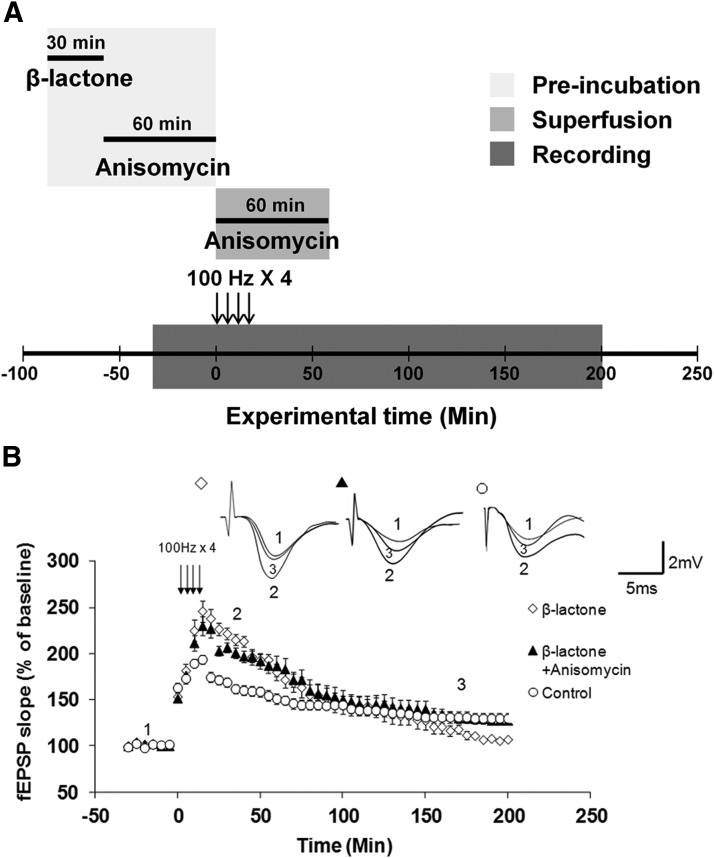
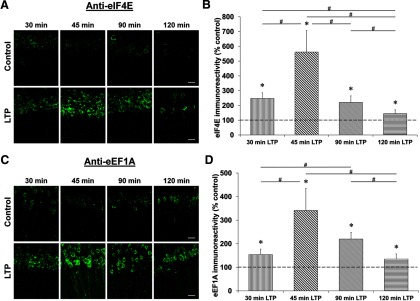
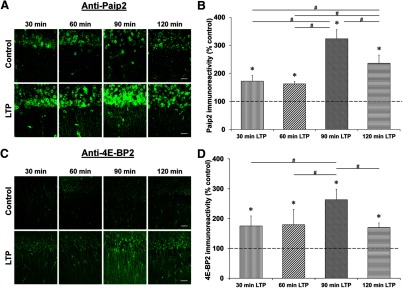
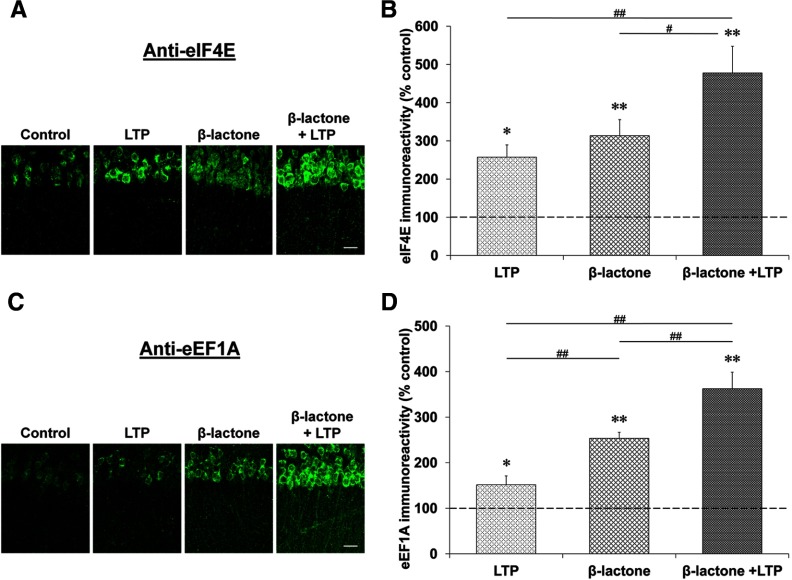
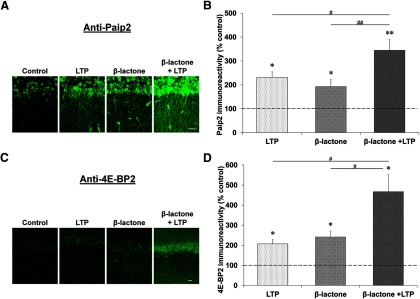
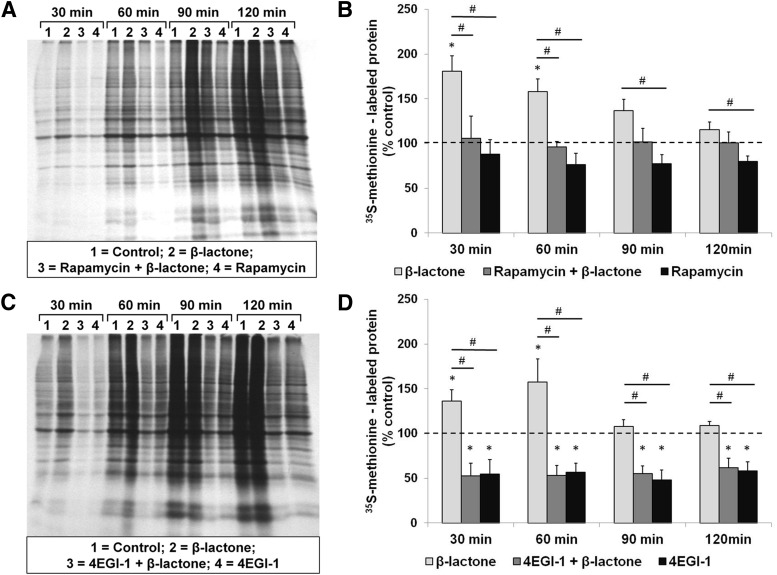
Proteolysis by the ubiquitin-proteasome pathway appears to have a complex role in synaptic plasticity, but its various functions remain to be elucidated. Using late phase long-term potentiation (L-LTP) in the hippocampus of the mouse as a model for long-term synaptic plasticity, we previously showed that inhibition of the proteasome enhances induction but blocks maintenance of L-LTP. In this study, we investigated the possible mechanisms by which proteasome inhibition has opposite effects on L-LTP induction and maintenance. Our results show that inhibiting phosphatidyl inositol-3 kinase or blocking the interaction between eukaryotic initiation factors 4E (eIF4E) and 4G (eIF4G) reduces the enhancement of L-LTP induction brought about by proteasome inhibition suggesting interplay between proteolysis and the signaling pathway mediated by mammalian target of rapamycin (mTOR). Also, proteasome inhibition leads to accumulation of translational activators in the mTOR pathway such as eIF4E and eukaryotic elongation factor 1A (eEF1A) early during L-LTP causing increased induction. Furthermore, inhibition of the proteasome causes a buildup of translational repressors, such as polyadenylate-binding protein interacting protein 2 (Paip2) and eukaryotic initiation factor 4E-binding protein 2 (4E-BP2), during late stages of L-LTP contributing to the blockade of L-LTP maintenance. Thus, the proteasome plays a critical role in regulating protein synthesis during L-LTP by tightly controlling translation. Our results provide novel mechanistic insights into the interplay between protein degradation and protein synthesis in long-term synaptic plasticity.
Keywords: activator; late-phase LTP; protein synthesis; proteolysis; repressor; ubiquitin.
Figures
Similar articles
-
Proteasome regulates the mediators of cytoplasmic polyadenylation signaling during late-phase long-term potentiation.Neurosci Lett. 2014 Nov 7;583:199-204. doi: 10.1016/j.neulet.2014.09.038. Epub 2014 Sep 28. Neurosci Lett. 2014. PMID: 25263789 Free PMC article.
-
Proteasome inhibition enhances the induction and impairs the maintenance of late-phase long-term potentiation.Learn Mem. 2008 Apr 25;15(5):335-47. doi: 10.1101/lm.984508. Print 2008 May. Learn Mem. 2008. PMID: 18441292 Free PMC article.
-
A rapamycin-sensitive signaling pathway contributes to long-term synaptic plasticity in the hippocampus.Proc Natl Acad Sci U S A. 2002 Jan 8;99(1):467-72. doi: 10.1073/pnas.012605299. Epub 2001 Dec 26. Proc Natl Acad Sci U S A. 2002. PMID: 11756682 Free PMC article.
-
BDNF mechanisms in late LTP formation: A synthesis and breakdown.Neuropharmacology. 2014 Jan;76 Pt C:664-76. doi: 10.1016/j.neuropharm.2013.06.024. Epub 2013 Jul 2. Neuropharmacology. 2014. PMID: 23831365 Review.
-
BDNF-induced local protein synthesis and synaptic plasticity.Neuropharmacology. 2014 Jan;76 Pt C:639-56. doi: 10.1016/j.neuropharm.2013.04.005. Epub 2013 Apr 16. Neuropharmacology. 2014. PMID: 23602987 Review.
Cited by
-
Dynamics of Hippocampal Protein Expression During Long-term Spatial Memory Formation.Mol Cell Proteomics. 2016 Feb;15(2):523-41. doi: 10.1074/mcp.M115.051318. Epub 2015 Nov 23. Mol Cell Proteomics. 2016. PMID: 26598641 Free PMC article.
-
Recent insights on principles of synaptic protein degradation.F1000Res. 2017 May 15;6:675. doi: 10.12688/f1000research.10599.1. eCollection 2017. F1000Res. 2017. PMID: 28620464 Free PMC article. Review.
-
The roles of protein expression in synaptic plasticity and memory consolidation.Front Mol Neurosci. 2014 Nov 12;7:86. doi: 10.3389/fnmol.2014.00086. eCollection 2014. Front Mol Neurosci. 2014. PMID: 25429258 Free PMC article. Review.
-
Perturbed proteostasis in autism spectrum disorders.J Neurochem. 2016 Dec;139(6):1081-1092. doi: 10.1111/jnc.13723. Epub 2016 Aug 4. J Neurochem. 2016. PMID: 27365114 Free PMC article. Review.
-
Proteolysis, synaptic plasticity and memory.Neurobiol Learn Mem. 2017 Feb;138:98-110. doi: 10.1016/j.nlm.2016.09.003. Epub 2016 Sep 7. Neurobiol Learn Mem. 2017. PMID: 27614141 Free PMC article. Review.
References
-
- Chain DG, Casadio A, Schacher S, Hegde AN, Valbrun M, Yamamoto N, Goldberg AL, Bartsch D, Kandel ER, Schwartz JH. Mechanisms for generating the autonomous cAMP-dependent protein kinase required for long-term facilitation in Aplysia. Neuron. 1999;22:147–156. doi: 10.1016/S0896-6273(00)80686-8. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous