

The multiple faces of leukocyte interstitial migration
- PMID: 24573488
- PMCID: PMC4118216
- DOI: 10.1007/s00281-014-0418-8
The multiple faces of leukocyte interstitial migration
Abstract
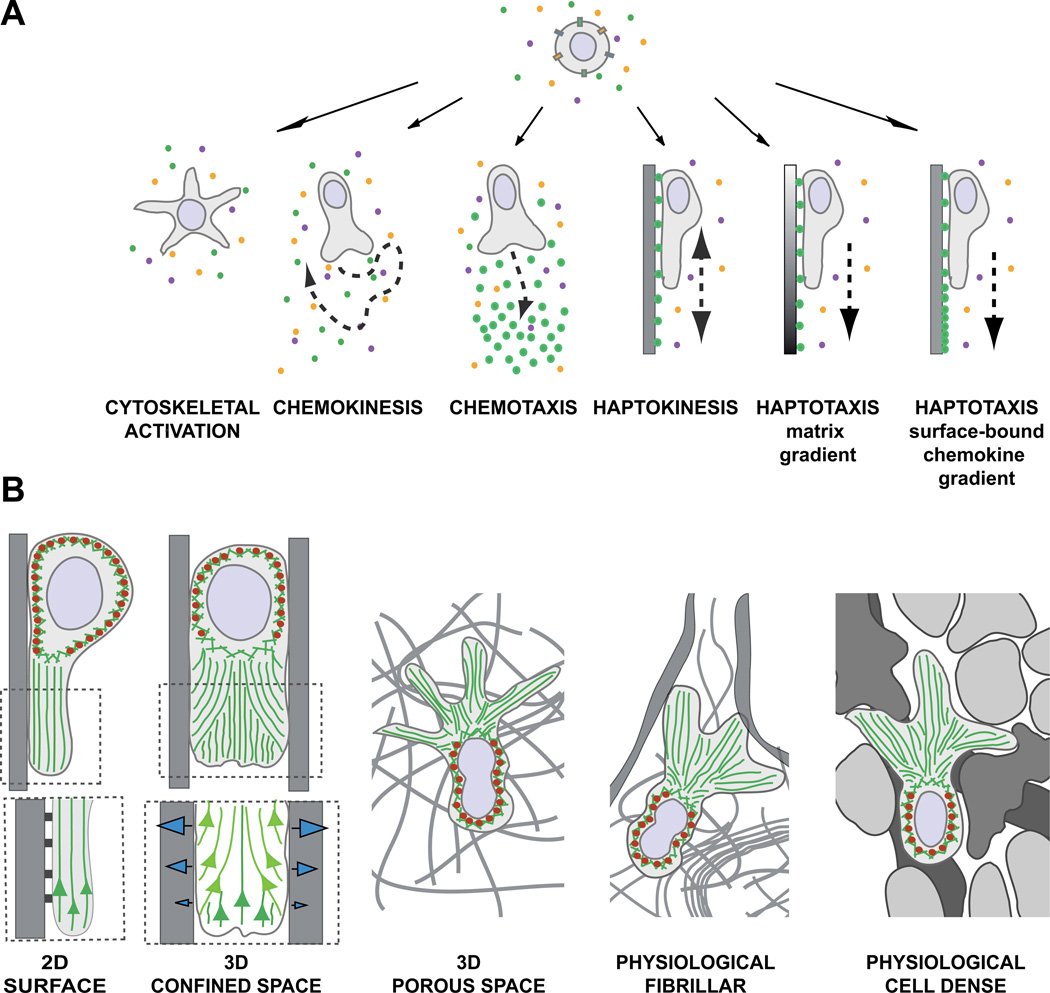
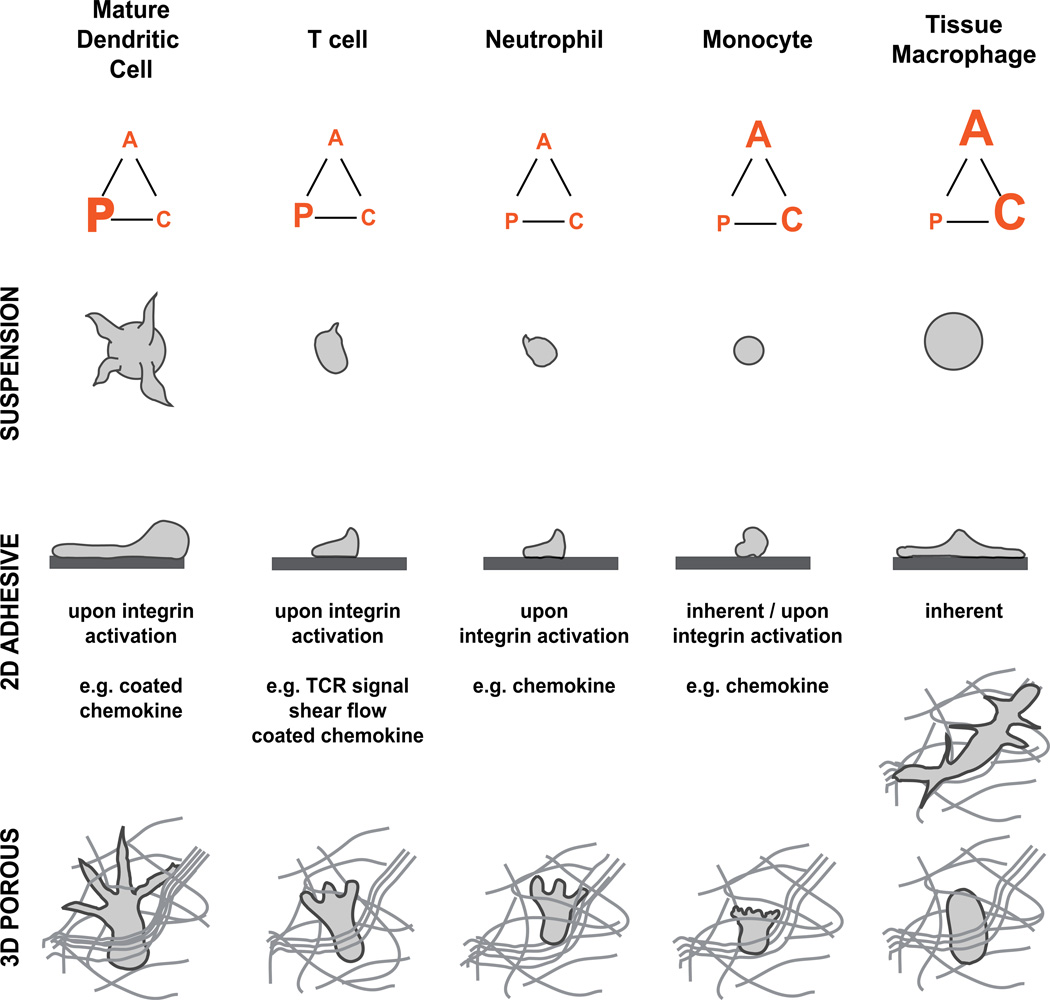
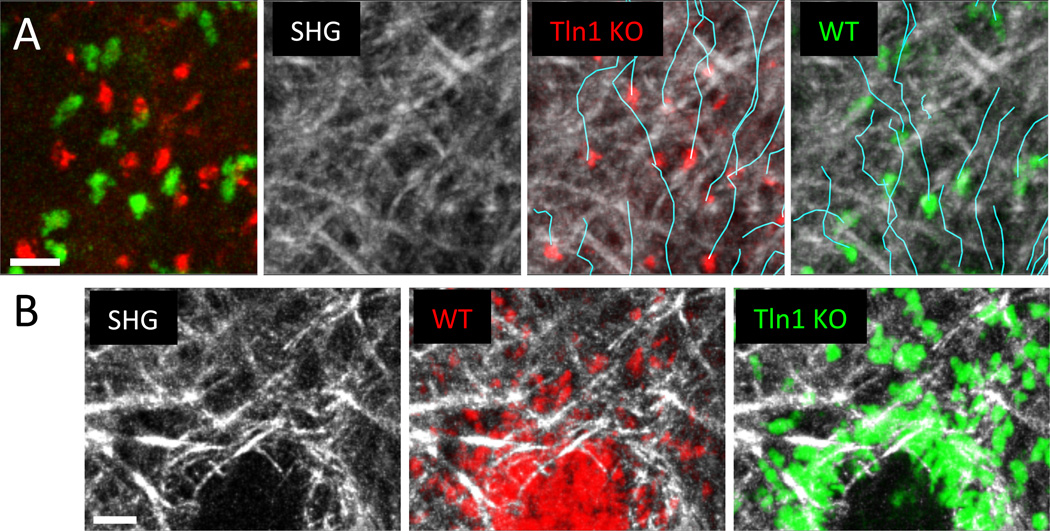
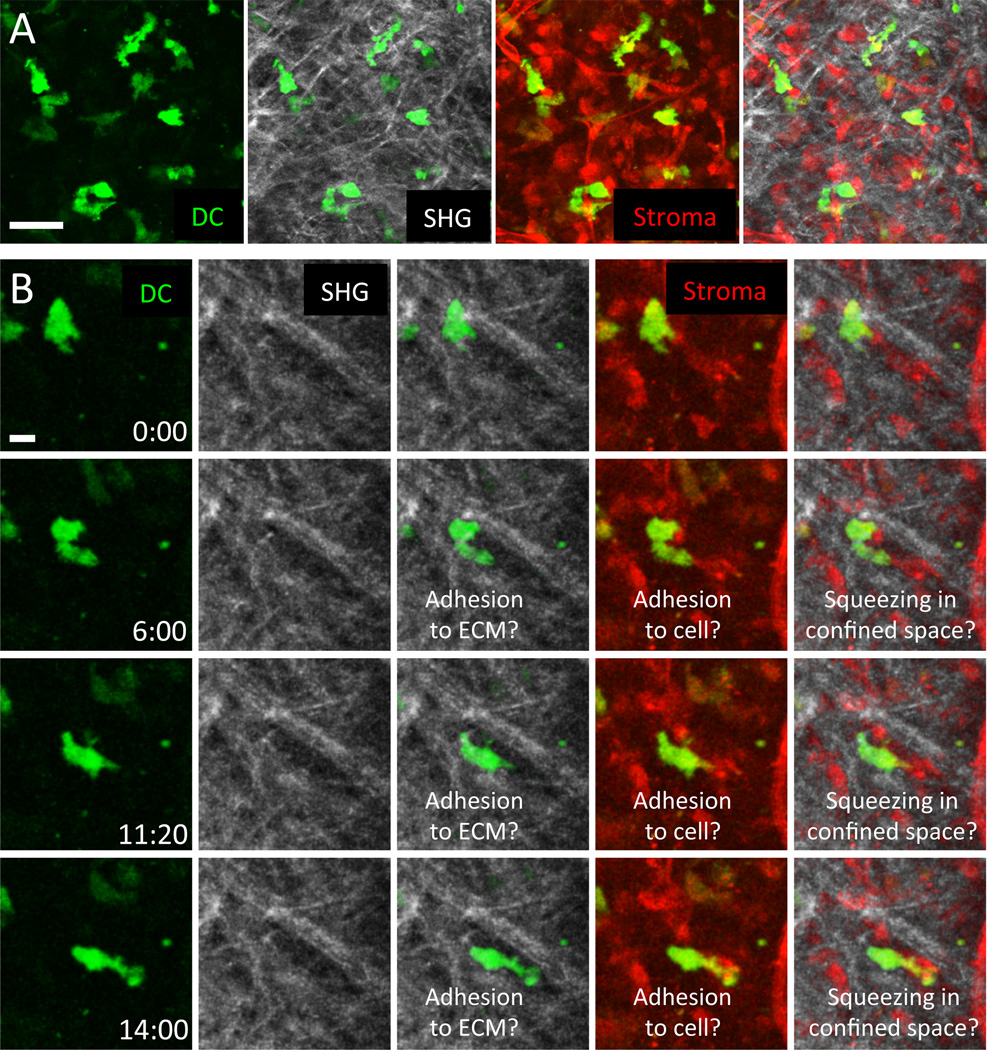
Spatiotemporal control of leukocyte dynamics within tissues is critical for successful innate and adaptive immune responses. Homeostatic trafficking and coordinated infiltration into and within sites of inflammation and infection rely on signaling in response to extracellular cues that in turn controls a variety of intracellular protein networks regulating leukocyte motility, migration, chemotaxis, positioning, and cell-cell interaction. In contrast to mesenchymal cells, leukocytes migrate in an amoeboid fashion by rapid cycles of actin polymerization and actomyosin contraction, and their migration in tissues is generally referred to as low adhesive and nonproteolytic. The interplay of actin network expansion, contraction, and adhesion shapes the exact mode of amoeboid migration, and in this review, we explore how leukocyte subsets potentially harness the same basic biomechanical mechanisms in a cell-type-specific manner. Most of our detailed understanding of these processes derives from in vitro migration studies in three-dimensional gels and confined spaces that mimic geometrical aspects of physiological tissues. We summarize these in vitro results and then critically compare them to data from intravital imaging of leukocyte interstitial migration in mouse tissues. We outline the technical challenges of obtaining conclusive mechanistic results from intravital studies, discuss leukocyte migration strategies in vivo, and present examples of mode switching during physiological interstitial migration. These findings are also placed in the context of leukocyte migration defects in primary immunodeficiencies. This overview of both in vitro and in vivo studies highlights recent progress in understanding the molecular and biophysical mechanisms that shape robust leukocyte migration responses in physiologically complex and heterogeneous environments.
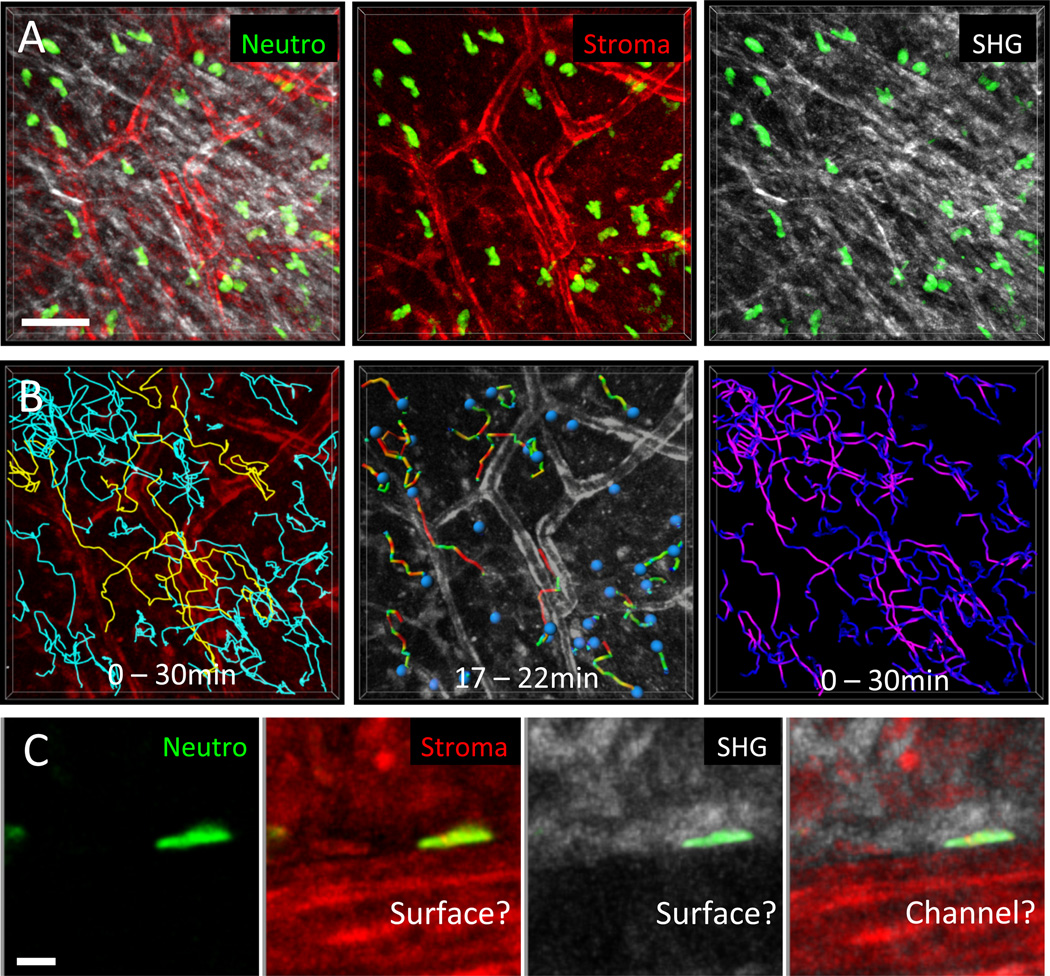
Figures
References
-
- von Andrian UH, Mempel TR. Homing and cellular traffic in lymph nodes. Nat Rev Immunol. 2003;3(11):867–878. - PubMed
-
- Luster AD, Alon R, von Andrian UH. Immune cell migration in inflammation: present and future therapeutic targets. Nat Immunol. 2005;6(12):1182–1190. - PubMed
-
- Friedl P, Weigelin B. Interstitial leukocyte migration and immune function. Nat Immunol. 2008;9(9):960–969. - PubMed
-
- Lämmermann T, Sixt M. Mechanical modes of ‘amoeboid’ cell migration. Curr Opin Cell Biol. 2009;21(5):636–644. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
