

Established and potential physiological roles of bicarbonate-sensing soluble adenylyl cyclase (sAC) in aquatic animals
- PMID: 24574382
- PMCID: PMC3936885
- DOI: 10.1242/jeb.086157
Established and potential physiological roles of bicarbonate-sensing soluble adenylyl cyclase (sAC) in aquatic animals
Abstract
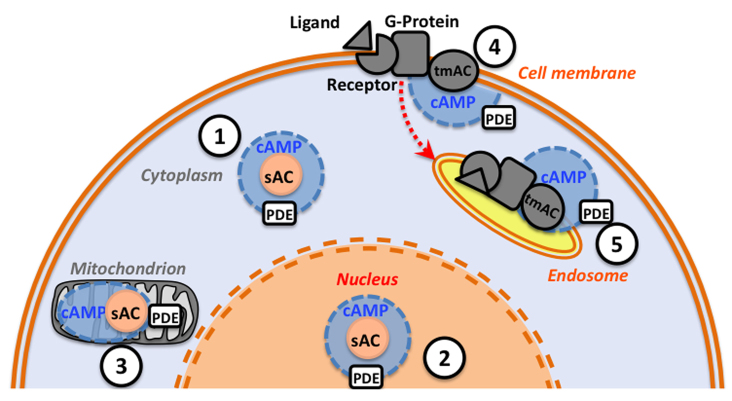
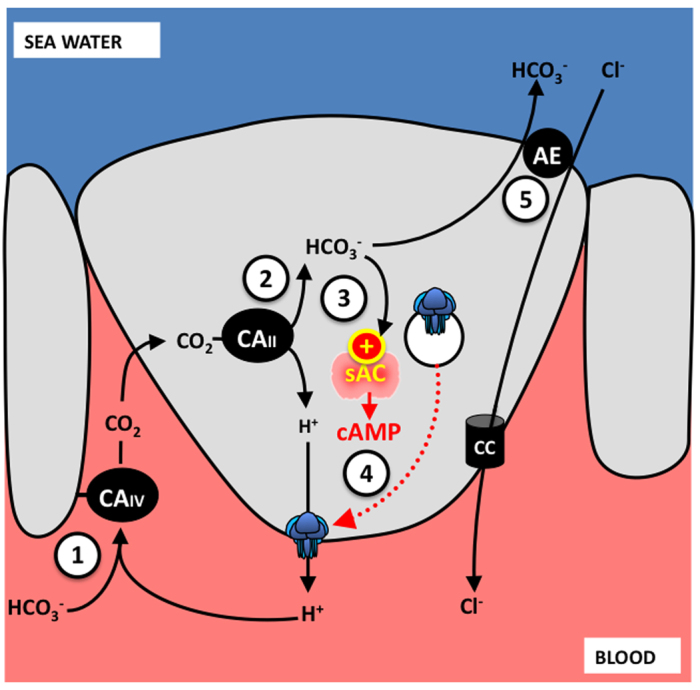
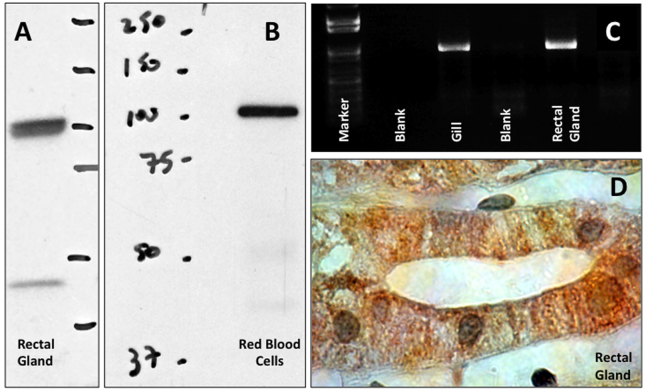
Soluble adenylyl cyclase (sAC) is a recently recognized source of the signaling molecule cyclic AMP (cAMP) that is genetically and biochemically distinct from the classic G-protein-regulated transmembrane adenylyl cyclases (tmACs). Mammalian sAC is distributed throughout the cytoplasm and it may be present in the nucleus and inside mitochondria. sAC activity is directly stimulated by HCO3(-), and sAC has been confirmed to be a HCO3(-) sensor in a variety of mammalian cell types. In addition, sAC can functionally associate with carbonic anhydrases to act as a de facto sensor of pH and CO2. The two catalytic domains of sAC are related to HCO3(-)-regulated adenylyl cyclases from cyanobacteria, suggesting the cAMP pathway is an evolutionarily conserved mechanism for sensing CO2 levels and/or acid/base conditions. Reports of sAC in aquatic animals are still limited but are rapidly accumulating. In shark gills, sAC senses blood alkalosis and triggers compensatory H(+) absorption. In the intestine of bony fishes, sAC modulates NaCl and water absorption. And in sea urchin sperm, sAC may participate in the initiation of flagellar movement and in the acrosome reaction. Bioinformatics and RT-PCR results reveal that sAC orthologs are present in most animal phyla. This review summarizes the current knowledge on the physiological roles of sAC in aquatic animals and suggests additional functions in which sAC may be involved.
Keywords: Acid/base; Carbonic anhydrase; Proton pump; V-ATPase; cAMP; pH sensing.
Figures

Similar articles
-
Pharmacological modulation of the CO2/HCO3-/pH-, calcium-, and ATP-sensing soluble adenylyl cyclase.Pharmacol Ther. 2018 Oct;190:173-186. doi: 10.1016/j.pharmthera.2018.05.008. Epub 2018 May 26. Pharmacol Ther. 2018. PMID: 29807057 Free PMC article. Review.
-
Bicarbonate-sensing soluble adenylyl cyclase is present in the cell cytoplasm and nucleus of multiple shark tissues.Physiol Rep. 2017 Jan;5(2):e13090. doi: 10.14814/phy2.13090. Physiol Rep. 2017. PMID: 28108644 Free PMC article.
-
Soluble adenylyl cyclase is an acid-base sensor in epithelial base-secreting cells.Am J Physiol Cell Physiol. 2016 Aug 1;311(2):C340-9. doi: 10.1152/ajpcell.00089.2016. Epub 2016 Jun 22. Am J Physiol Cell Physiol. 2016. PMID: 27335168 Free PMC article.
-
Bicarbonate-regulated soluble adenylyl cyclase.JOP. 2001 Jul;2(4 Suppl):154-8. JOP. 2001. PMID: 11875252 Review.
-
Bicarbonate-sensing soluble adenylyl cyclase is an essential sensor for acid/base homeostasis.Proc Natl Acad Sci U S A. 2010 Jan 5;107(1):442-7. doi: 10.1073/pnas.0911790107. Epub 2009 Dec 14. Proc Natl Acad Sci U S A. 2010. PMID: 20018667 Free PMC article.
Cited by
-
Subcellular localization and characterization of estrogenic pathway regulators and mediators in Atlantic salmon spermatozoal cells.Histochem Cell Biol. 2018 Jan;149(1):75-96. doi: 10.1007/s00418-017-1611-3. Epub 2017 Oct 5. Histochem Cell Biol. 2018. PMID: 28983690
-
Comparative physiology reveals heat stress disrupts acid-base homeostasis independent of symbiotic state in the model cnidarian Exaiptasia diaphana.J Exp Biol. 2024 Feb 15;227(4):jeb246222. doi: 10.1242/jeb.246222. Epub 2024 Feb 22. J Exp Biol. 2024. PMID: 38269486 Free PMC article.
-
The silent loss of cell physiology hampers marine biosciences.PLoS Biol. 2022 May 12;20(5):e3001641. doi: 10.1371/journal.pbio.3001641. eCollection 2022 May. PLoS Biol. 2022. PMID: 35550624 Free PMC article.
-
Pharmacological modulation of the CO2/HCO3-/pH-, calcium-, and ATP-sensing soluble adenylyl cyclase.Pharmacol Ther. 2018 Oct;190:173-186. doi: 10.1016/j.pharmthera.2018.05.008. Epub 2018 May 26. Pharmacol Ther. 2018. PMID: 29807057 Free PMC article. Review.
-
Gymnocypris przewalskii decreases cytosolic carbonic anhydrase expression to compensate for respiratory alkalosis and osmoregulation in the saline-alkaline lake Qinghai.J Comp Physiol B. 2016 Jan;186(1):83-95. doi: 10.1007/s00360-015-0939-z. J Comp Physiol B. 2016. PMID: 26475604
References
-
- Appukuttan A., Kasseckert S. A., Micoogullari M., Flacke J.-P., Kumar S., Woste A., Abdallah Y., Pott L., Reusch H. P., Ladilov Y. (2012). Type 10 adenylyl cyclase mediates mitochondrial Bax translocation and apoptosis of adult rat cardiomyocytes under simulated ischaemia/reperfusion. Cardiovasc. Res. 93, 340-349 - PubMed
-
- Barron M. E., Roa J. N. B., Tresguerres M. (2012). Pacific oyster mantle, gill and hemocytes express the bicarbonate-sensing enzyme soluble adenylyl cyclase. FASEB J. 26, 1070.2
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
