

Functional analysis of nuclear localization signals in VP1-2 homologues from all herpesvirus subfamilies
- PMID: 24574406
- PMCID: PMC4019078
- DOI: 10.1128/JVI.03797-13
Functional analysis of nuclear localization signals in VP1-2 homologues from all herpesvirus subfamilies
Abstract
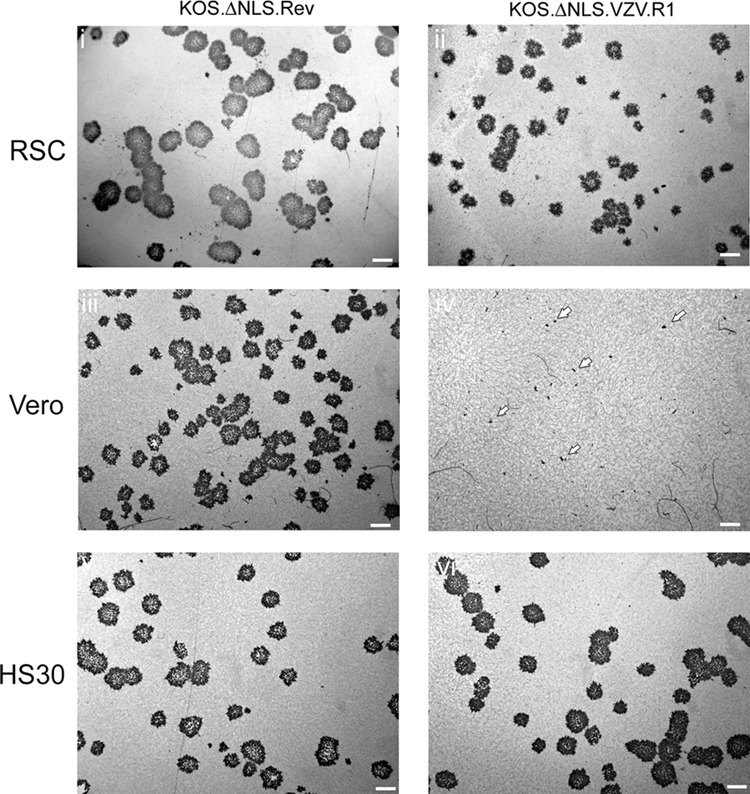
The herpes simplex virus (HSV) tegument protein VP1-2 contains an N-terminal nuclear localization signal (NLS) that is critical for capsid routing to the nuclear pore. Here we analyzed positionally conserved determinants in VP1-2 homologues from each of the alpha, beta, and gamma classes of human herpesviruses. The overall architectures of the VP1-2s were similar, with a conserved N-terminal ubiquitin-specific protease domain separated from an internal region by a linker that was quite poorly conserved in length and sequence. Within this linker region all herpesviruses contained a conserved, highly basic motif which nevertheless exhibited distinct class-specific features. The motif in HSV functioned as a monopartite NLS, while in varicella-zoster virus (VZV) activity required an adjacent basic section defining the motif as a bipartite NLS. Neither the beta- nor gammaherpesvirus VP1-2 motifs were identified by prediction algorithms, but they nevertheless functioned as efficient NLS motifs both in heterologous transfer assays and in HSV VP1-2. Furthermore, though with different efficiencies and with the exception of human herpesvirus 8 (HHV-8), these chimeric variants rescued the replication defect of an HSV mutant lacking its NLS motif. We demonstrate that the lysine at position 428 of HSV is critical for replication, with a single alanine substitution being sufficient to abrogate NLS function and virus growth. We conclude that the basic motifs of each of the VP1-2 proteins are likely to confer a similar function in capsid entry in the homologous setting and that while there is flexibility in the exact type of motif employed, specific individual residues are critical for function.
Importance: To successfully infect cells, all herpesviruses, along with many other viruses, e.g., HIV, hepatitis B virus, and influenza virus, must navigate through the cytoplasmic environment and dock with nuclear pores for transport of their genomes into the nucleus. However, we still have a limited understanding of the detailed mechanisms involved. Insight into these events is needed and could offer opportunities for therapeutic intervention. This work investigated the role of a specific determinant in the structural protein VP1-2 in herpesvirus entry. We examined this determinant in representative VP1-2s from all herpesvirus subfamilies, demonstrated NLS function, dissected key residues, and showed functional relevance in rescuing replication of the mutant blocked in capsid navigation to the pore. The results are important and strongly support our conclusions of the generality that these motifs are crucial for entry of all herpesviruses. They also facilitate future analysis on selective host interactions and possible routes to disrupt function.
Figures









Similar articles
-
Comprehensive Analysis of the Tegument Proteins Involved in Capsid Transport and Virion Morphogenesis of Alpha, Beta and Gamma Herpesviruses.Viruses. 2023 Oct 6;15(10):2058. doi: 10.3390/v15102058. Viruses. 2023. PMID: 37896835 Free PMC article. Review.
-
The role of nuclear localization signal in parvovirus life cycle.Virol J. 2017 Apr 14;14(1):80. doi: 10.1186/s12985-017-0745-1. Virol J. 2017. PMID: 28410597 Free PMC article. Review.
-
A Nuclear localization signal in herpesvirus protein VP1-2 is essential for infection via capsid routing to the nuclear pore.J Virol. 2012 Sep;86(17):8998-9014. doi: 10.1128/JVI.01209-12. Epub 2012 Jun 20. J Virol. 2012. PMID: 22718835 Free PMC article.
-
Identification of a highly conserved, functional nuclear localization signal within the N-terminal region of herpes simplex virus type 1 VP1-2 tegument protein.J Virol. 2008 Jun;82(11):5234-44. doi: 10.1128/JVI.02497-07. Epub 2008 Apr 2. J Virol. 2008. PMID: 18385239 Free PMC article.
-
Classic nuclear localization signals and a novel nuclear localization motif are required for nuclear transport of porcine parvovirus capsid proteins.J Virol. 2014 Oct;88(20):11748-59. doi: 10.1128/JVI.01717-14. Epub 2014 Jul 30. J Virol. 2014. PMID: 25078698 Free PMC article.
Cited by
-
Comprehensive Analysis of the Tegument Proteins Involved in Capsid Transport and Virion Morphogenesis of Alpha, Beta and Gamma Herpesviruses.Viruses. 2023 Oct 6;15(10):2058. doi: 10.3390/v15102058. Viruses. 2023. PMID: 37896835 Free PMC article. Review.
-
pUL36 Deubiquitinase Activity Augments Both the Initiation and the Progression of Lytic Herpes Simplex Virus Infection in IFN-Primed Cells.J Virol. 2022 Nov 23;96(22):e0096322. doi: 10.1128/jvi.00963-22. Epub 2022 Oct 31. J Virol. 2022. PMID: 36314822 Free PMC article.
-
Involvement of the N-Terminal Deubiquitinating Protease Domain of Human Cytomegalovirus UL48 Tegument Protein in Autoubiquitination, Virion Stability, and Virus Entry.J Virol. 2016 Jan 13;90(6):3229-42. doi: 10.1128/JVI.02766-15. J Virol. 2016. PMID: 26764006 Free PMC article.
-
The role of nuclear localization signal in parvovirus life cycle.Virol J. 2017 Apr 14;14(1):80. doi: 10.1186/s12985-017-0745-1. Virol J. 2017. PMID: 28410597 Free PMC article. Review.
-
Conserved Tryptophan Motifs in the Large Tegument Protein pUL36 Are Required for Efficient Secondary Envelopment of Herpes Simplex Virus Capsids.J Virol. 2016 May 12;90(11):5368-5383. doi: 10.1128/JVI.03167-15. Print 2016 Jun 1. J Virol. 2016. PMID: 27009950 Free PMC article.
References
-
- Flint SJ, Enquist LW, Krug RM, Racaniello VR, Skalka AM. 2009. Principles of virology. ASM Press, Washington, DC
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
