

Functional divergence between the two P1-P2 stalk dimers on the ribosome in their interaction with ricin A chain
- PMID: 24576056
- PMCID: PMC4100481
- DOI: 10.1042/BJ20140014
Functional divergence between the two P1-P2 stalk dimers on the ribosome in their interaction with ricin A chain
Abstract
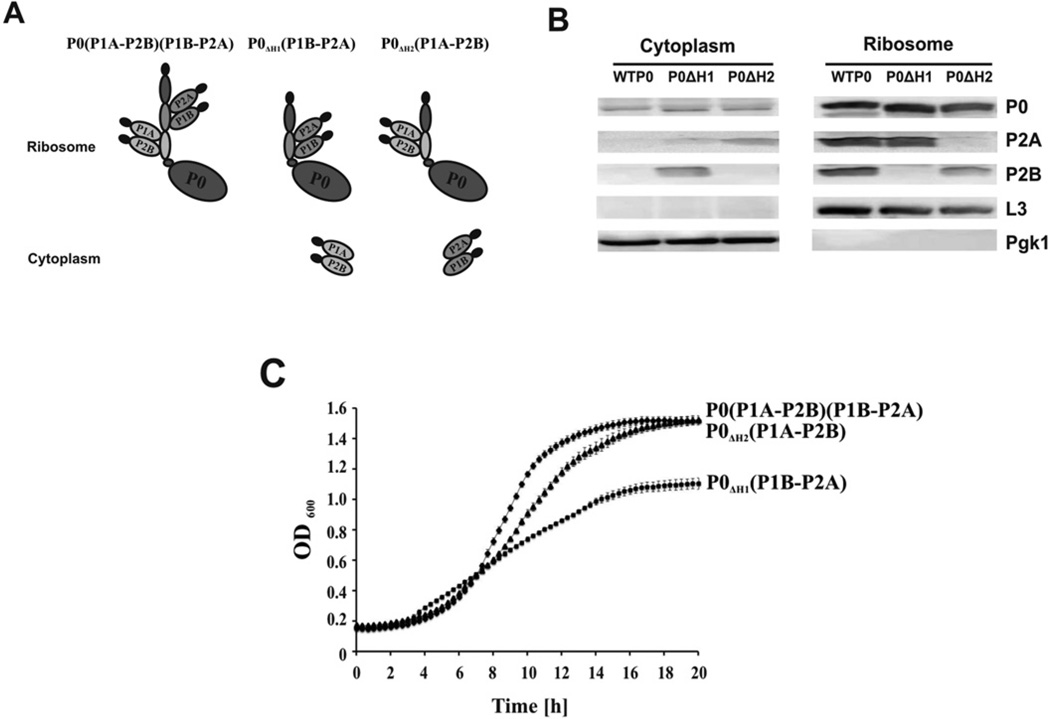
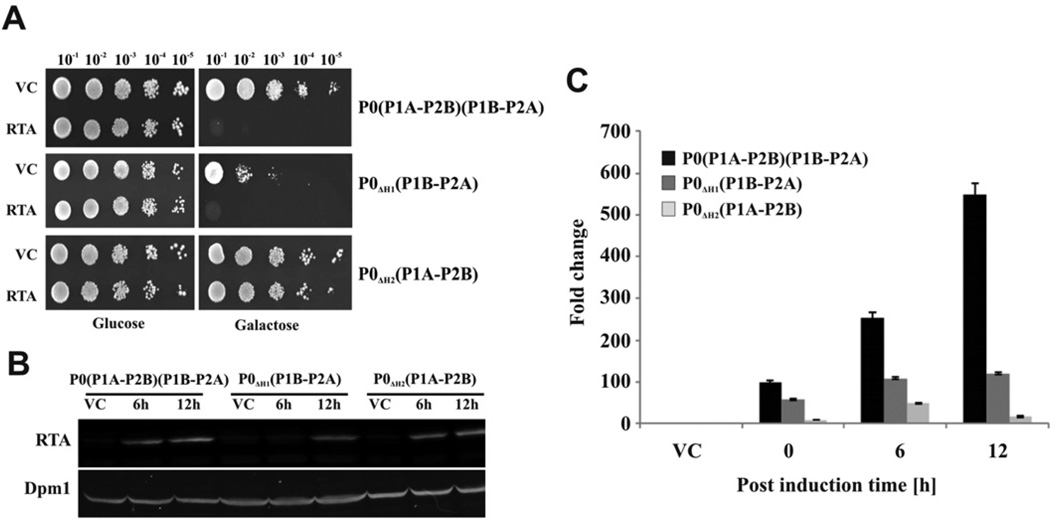
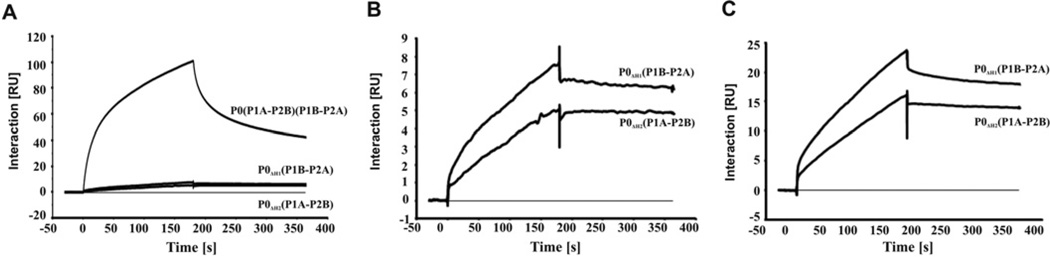
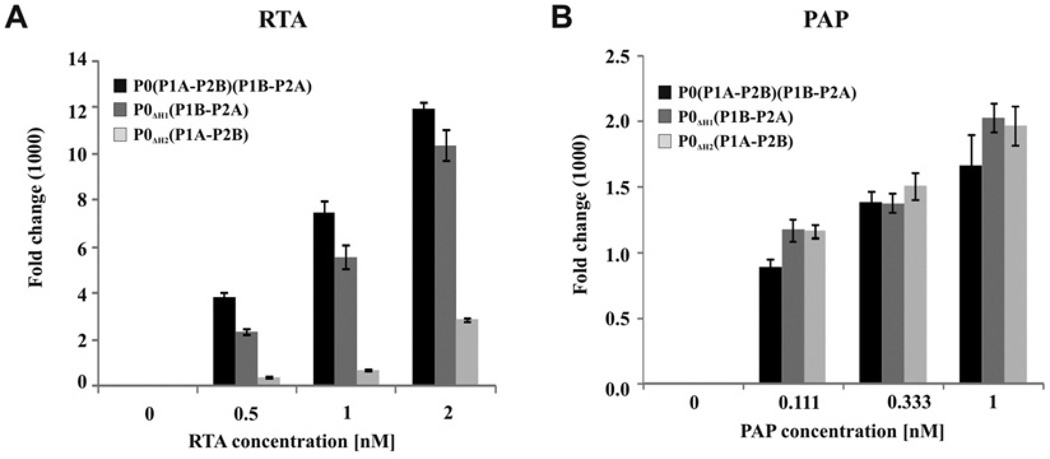
The eukaryotic stalk, which is responsible for the recruitment of translation factors, is a pentamer containing two P1-P2 dimers with unclear modes of action. In Saccharomyces cerevisiae, P1/P2 proteins (individual P1 and P2 proteins) are organized into two distinct dimers, P1A-P2B and P1B-P2A. To investigate the functional contribution of each dimer on the ribosome, RTA (ricin A chain), which binds to the stalk to depurinate the SRL (sarcin/ricin loop), was used as a molecular probe in yeast mutants in which the binding site for one or the other dimer on P0 was deleted. Ribosome depurination and toxicity of RTA were greatly reduced in mutants containing only P1A-P2B on the ribosome, whereas those with only P1B-P2A were reduced less in depurination and were unaffected in toxicity. Ribosomes bearing P1B-P2A were depurinated by RTA at a similar level as wild-type, but ribosomes bearing P1A-P2B were depurinated at a much lower level in vitro. The latter ribosomes showed the lowest association and almost no dissociation with RTA by surface plasmon resonance. These results indicate that the P1B-P2A dimer is more critical for facilitating the access of RTA to the SRL, providing the first in vivo evidence for functional divergence between the two stalk dimers on the ribosome.
Figures
Similar articles
-
Human ribosomal P1-P2 heterodimer represents an optimal docking site for ricin A chain with a prominent role for P1 C-terminus.Sci Rep. 2017 Jul 17;7(1):5608. doi: 10.1038/s41598-017-05675-5. Sci Rep. 2017. PMID: 28717148 Free PMC article.
-
The P1/P2 proteins of the human ribosomal stalk are required for ribosome binding and depurination by ricin in human cells.FEBS J. 2012 Oct;279(20):3925-36. doi: 10.1111/j.1742-4658.2012.08752.x. Epub 2012 Sep 11. FEBS J. 2012. PMID: 22909382 Free PMC article.
-
The ribosomal stalk is required for ribosome binding, depurination of the rRNA and cytotoxicity of ricin A chain in Saccharomyces cerevisiae.Mol Microbiol. 2008 Dec;70(6):1441-52. doi: 10.1111/j.1365-2958.2008.06492.x. Epub 2008 Oct 30. Mol Microbiol. 2008. PMID: 19019145 Free PMC article.
-
Interaction of ricin and Shiga toxins with ribosomes.Curr Top Microbiol Immunol. 2012;357:1-18. doi: 10.1007/82_2011_174. Curr Top Microbiol Immunol. 2012. PMID: 21910078 Free PMC article. Review.
-
Proteins P1, P2, and P0, components of the eukaryotic ribosome stalk. New structural and functional aspects.Biochem Cell Biol. 1995 Nov-Dec;73(11-12):959-68. doi: 10.1139/o95-103. Biochem Cell Biol. 1995. PMID: 8722011 Review.
Cited by
-
Conserved Arginines at the P-Protein Stalk Binding Site and the Active Site Are Critical for Ribosome Interactions of Shiga Toxins but Do Not Contribute to Differences in the Affinity of the A1 Subunits for the Ribosome.Infect Immun. 2016 Nov 18;84(12):3290-3301. doi: 10.1128/IAI.00630-16. Print 2016 Dec. Infect Immun. 2016. PMID: 27600507 Free PMC article.
-
Functional Assays for Measuring the Catalytic Activity of Ribosome Inactivating Proteins.Toxins (Basel). 2018 Jun 14;10(6):240. doi: 10.3390/toxins10060240. Toxins (Basel). 2018. PMID: 29899209 Free PMC article. Review.
-
How Ricin Damages the Ribosome.Toxins (Basel). 2019 Apr 27;11(5):241. doi: 10.3390/toxins11050241. Toxins (Basel). 2019. PMID: 31035546 Free PMC article. Review.
-
Human ribosomal P1-P2 heterodimer represents an optimal docking site for ricin A chain with a prominent role for P1 C-terminus.Sci Rep. 2017 Jul 17;7(1):5608. doi: 10.1038/s41598-017-05675-5. Sci Rep. 2017. PMID: 28717148 Free PMC article.
-
Intracellular Transport and Cytotoxicity of the Protein Toxin Ricin.Toxins (Basel). 2019 Jun 18;11(6):350. doi: 10.3390/toxins11060350. Toxins (Basel). 2019. PMID: 31216687 Free PMC article. Review.
References
-
- Liljas A, Ehrenberg M. Structural Aspects of Protein Synthesis. 2nd Edition. Singapore: World Scientific Publishing; 2012. pp. 1–492.
-
- Helgstrand M, Mandava CS, Mulder FA, Liljas A, Sanyal S, Akke M. The ribosomal stalk binds to translation factors IF2, EF-Tu, EF-G and RF3 via a conserved region of the L12 C-terminal domain. J. Mol. Biol. 2007;365:468–479. - PubMed
-
- Nomura N, Honda T, Baba K, Naganuma T, Tanzawa T, Arisaka F, Noda M, Uchiyama S, Tanaka I, Yao M, et al. Archaeal ribosomal stalk protein interacts with translation factors in a nucleotide-independent manner via its conserved C terminus. Proc. Natl. Acad. Sci. U.S.A. 2012;109:3748–3753. - PMC - PubMed
-
- Diaconu M, Kothe U, Schlunzen F, Fischer N, Harms JM, Tonevitsky AG, Stark H, Rodnina MV, Wahl MC. Structural basis for the function of the ribosomal L7/12 stalk in factor binding and GTPase activation. Cell. 2005;121:991–1004. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous