

New insights into FAK function and regulation during spermatogenesis
- PMID: 24578181
- PMCID: PMC4139008
- DOI: 10.14670/HH-29.977
New insights into FAK function and regulation during spermatogenesis
Abstract
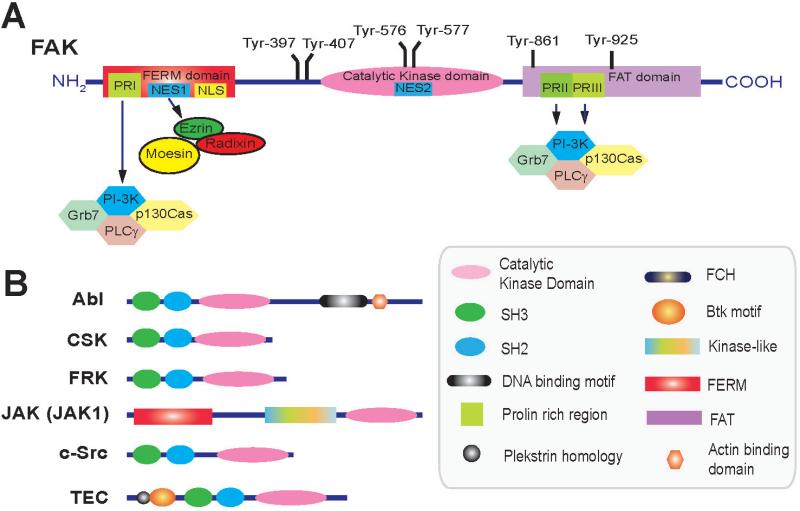
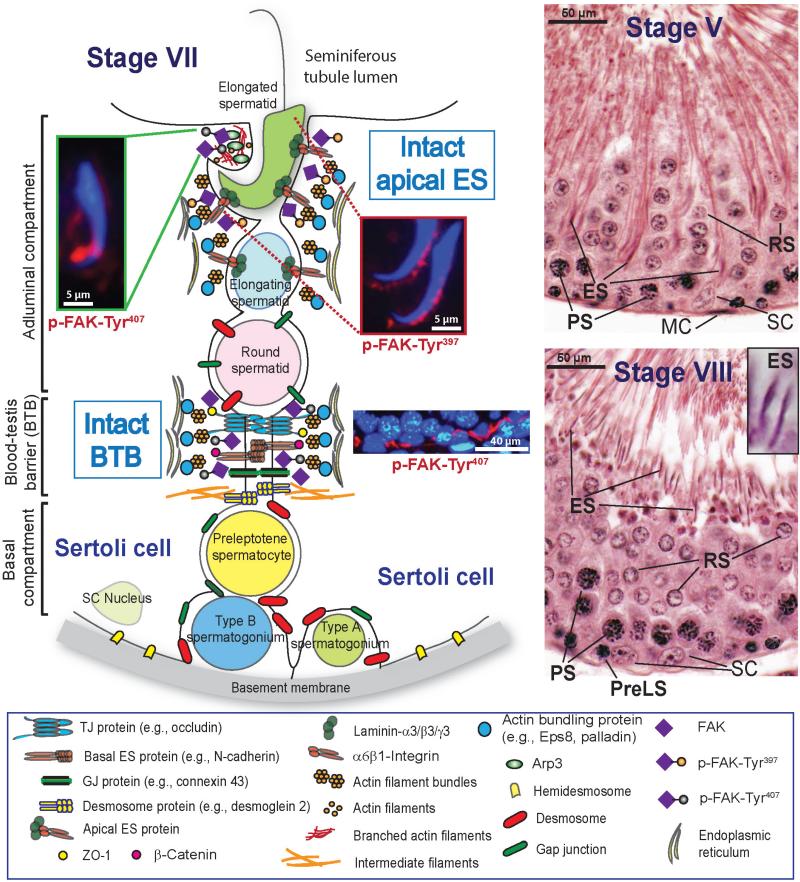
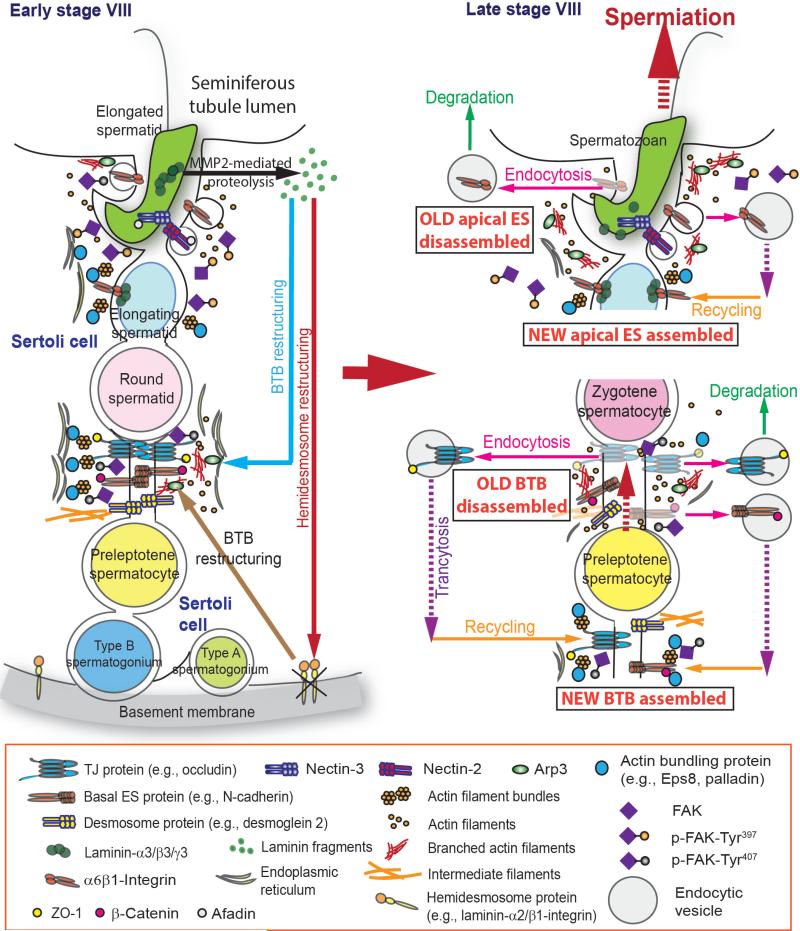
Germ cell transport across the seminiferous epithelium during the epithelial cycle is crucial to spermatogenesis, although molecular mechanism(s) that regulate these events remain unknown. Studies have shown that spatiotemporal expression of crucial regulatory proteins during the epithelial cycle represents an efficient and physiologically important mechanism to regulate spermatogenesis without involving de novo synthesis of proteins and/or expression of genes. Herein, we critically review the role of focal adhesion kinase (FAK) in coordinating the transport of spermatids and preleptotene spermatocytes across the epithelium and the BTB, respectively, along the apical ectoplasmic specialization (ES) - blood-testis barrier - basement membrane (BM) functional axis during spermatogenesis. In the testis, p-FAK-Tyr³⁸⁷ and p-FAK-Tyr⁴⁰⁷ are spatiotemporally expressed during the epithelial cycle at the actin-rich anchoring junction known as ES, regulating cell adhesion at the Sertoli-spermatid (apical ES) and Sertoli cell-cell (basal ES) interface. Phosphorylated forms of FAK exert their effects by regulating the homeostasis of F-actin at the ES, mediated via their effects on actin polymerization so that microfilaments are efficiently re-organized, such as from their "bundled" to "de-bundled/branched" configuration and vice versa during the epithelial cycle to facilitate the transport of: (i) spermatids across the epithelium, and (ii) preleptotene spermatocytes across the BTB. In summary, p-FAK-Tyr⁴⁰⁷ and p-FAK-Tyr³⁸⁷ are important regulators of spermatogenesis which serve as molecular switches that turn "on" and "off" adhesion function at the apical ES and the basal ES/BTB, mediated via their spatiotemporal expression during the epithelial cycle. A hypothetical model depicting the role of these two molecular switches is also proposed.
Figures
Similar articles
-
Focal adhesion kinase is a regulator of F-actin dynamics: New insights from studies in the testis.Spermatogenesis. 2013 Jul 1;3(3):e25385. doi: 10.4161/spmg.25385. Epub 2013 Jun 21. Spermatogenesis. 2013. PMID: 24381802 Free PMC article. Review.
-
Actin-bundling protein plastin 3 is a regulator of ectoplasmic specialization dynamics during spermatogenesis in the rat testis.FASEB J. 2015 Sep;29(9):3788-805. doi: 10.1096/fj.14-267997. Epub 2015 Jun 5. FASEB J. 2015. PMID: 26048141 Free PMC article.
-
Focal adhesion kinase and actin regulatory/binding proteins that modulate F-actin organization at the tissue barrier: Lesson from the testis.Tissue Barriers. 2013 Apr 1;1(2):e24252. doi: 10.4161/tisb.24252. Tissue Barriers. 2013. PMID: 24665388 Free PMC article.
-
p-FAK-Tyr(397) regulates spermatid adhesion in the rat testis via its effects on F-actin organization at the ectoplasmic specialization.Am J Physiol Endocrinol Metab. 2013 Sep 15;305(6):E687-99. doi: 10.1152/ajpendo.00254.2013. Epub 2013 Jul 23. Am J Physiol Endocrinol Metab. 2013. PMID: 23880313 Free PMC article.
-
Cell polarity proteins and spermatogenesis.Semin Cell Dev Biol. 2016 Nov;59:62-70. doi: 10.1016/j.semcdb.2016.06.008. Epub 2016 Jun 9. Semin Cell Dev Biol. 2016. PMID: 27292315 Free PMC article. Review.
Cited by
-
Design, Synthesis, and Evaluation of Highly Potent FAK-Targeting PROTACs.ACS Med Chem Lett. 2019 Oct 22;11(10):1855-1862. doi: 10.1021/acsmedchemlett.9b00372. eCollection 2020 Oct 8. ACS Med Chem Lett. 2019. PMID: 33062164 Free PMC article.
-
Protein degradation technology: a strategic paradigm shift in drug discovery.J Hematol Oncol. 2021 Sep 6;14(1):138. doi: 10.1186/s13045-021-01146-7. J Hematol Oncol. 2021. PMID: 34488823 Free PMC article. Review.
-
PROTACs: great opportunities for academia and industry.Signal Transduct Target Ther. 2019 Dec 24;4:64. doi: 10.1038/s41392-019-0101-6. eCollection 2019. Signal Transduct Target Ther. 2019. PMID: 31885879 Free PMC article. Review.
-
MiRNAs Expression Profiling of Bovine (Bos taurus) Testes and Effect of bta-miR-146b on Proliferation and Apoptosis in Bovine Male Germline Stem Cells.Int J Mol Sci. 2020 May 28;21(11):3846. doi: 10.3390/ijms21113846. Int J Mol Sci. 2020. PMID: 32481702 Free PMC article.
-
Male germline recombination of a conditional allele by the widely used Dermo1-cre (Twist2-cre) transgene.Genesis. 2017 Sep;55(9):10.1002/dvg.23048. doi: 10.1002/dvg.23048. Epub 2017 Aug 14. Genesis. 2017. PMID: 28722198 Free PMC article.
References
-
- Amann RP, Howards SS. Daily spermatozoal production and epididymal spermatozoal reserves of the human male. J. Urol. 1980;124:211–215. - PubMed
-
- Ammoun S, Provenzano L, Zhou L, Barczyk M, Evans K, Hilton DA, Hafizi S, Hanemann CO. Axl/gas6/nfÎb signalling in schwannoma pathological proliferation, adhesion and survival. Oncogene. 2014;33:336–346. - PubMed
-
- André E, Becker-Andre M. Expression of an N-terminally truncated form of human focal adhesion kinase in brain. Biochem. Biophys. Res. Commun. 1993;190:140–147. - PubMed
-
- Bartles J, Wierda A, Zheng L. Identification and characterization of espin, an actin-binding protein localized to the f-actin-rich junctional plaques of sertoli cell ectoplasmic specializations. J. Cell Sci. 1996;109:1229–1239. - PubMed
-
- Beardsley A, Robertson DM, O'Donnell L. A complex containing α6β1-integrin and phosphorylated focal adhesion kinase between sertoli cells and elongated spermatids during spermatid release from the seminiferous epithelium. J. Endocrinol. 2006;190:759–770. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
