

Activation of PI3K and R-Ras signaling promotes the extension of sensory axons on inhibitory chondroitin sulfate proteoglycans
- PMID: 24578264
- PMCID: PMC4235675
- DOI: 10.1002/dneu.22174
Activation of PI3K and R-Ras signaling promotes the extension of sensory axons on inhibitory chondroitin sulfate proteoglycans
Abstract
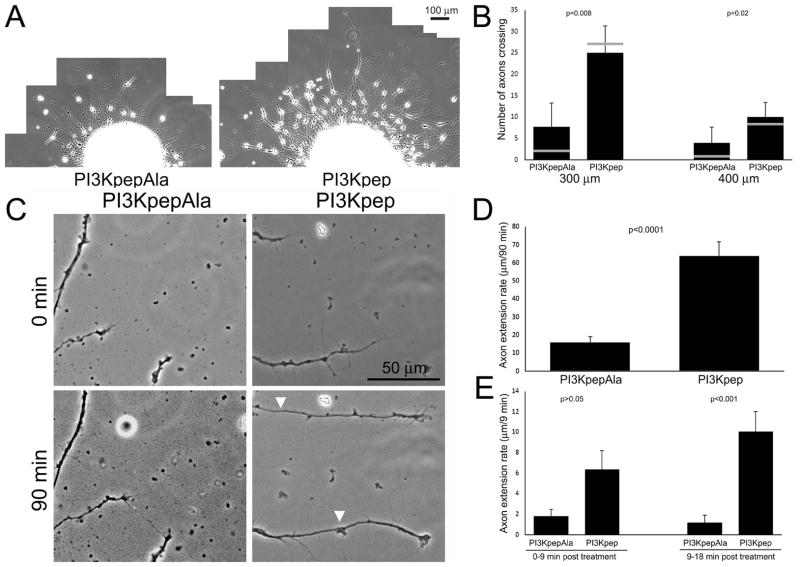
Chondroitin sulfate proteoglycans (CSPGs) are extracellular inhibitors of axon extension and plasticity, and cause growth cones to exhibit dystrophic behaviors. Phosphoinositide 3-kinase (PI3K) is a lipid kinase activated by axon growth promoting signals. In this study, we used embryonic chicken dorsal root ganglion neurons to determine if CSPGs impair signaling through PI3K. We report that CSPGs inhibit PI3K signaling in axons and growth cones, as evidenced by decreased levels of phosphorylated downstream kinases (Akt and S6). Direct activation of PI3K signaling, using a cell permeable phosphopeptide (PI3Kpep), countered the effects of CSPGs on growth cones and axon extension. Both overnight and acute treatment with PI3Kpep promoted axon extension on CSPG-coated substrates. The R-Ras GTPase is an upstream positive regulator of PI3K signaling. Expression of constitutively active R-Ras promoted axon extension and growth cone elaboration on CSPGs and permissive substrata. In contrast, an N-terminus-deleted constitutively active R-Ras, deficient in PI3K activation, promoted axon extension but not growth cone elaboration on CSPGs and permissive substrata. These data indicate that activation of R-Ras-PI3K signaling may be a viable approach for manipulating axon extension on CSPGs.
Keywords: Akt; S6; growth cone; neurite; regeneration.
© 2014 Wiley Periodicals, Inc.
Figures






References
-
- Bartus K, James ND, Bosch KD, Bradbury EJ. Chondroitin sulphate proteoglycans: key modulators of spinal cord and brain plasticity. Exp Neurol. 2012;235:5–17. - PubMed
-
- Carriere A, Ray H, Blenis J, Roux PP. The RSK factors of activating the Ras/MAPK signaling cascade. Front Biosci. 2008;13:4258–75. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
