

Promoter-bound trinucleotide repeat mRNA drives epigenetic silencing in fragile X syndrome
- PMID: 24578575
- PMCID: PMC4357282
- DOI: 10.1126/science.1245831
Promoter-bound trinucleotide repeat mRNA drives epigenetic silencing in fragile X syndrome
Abstract
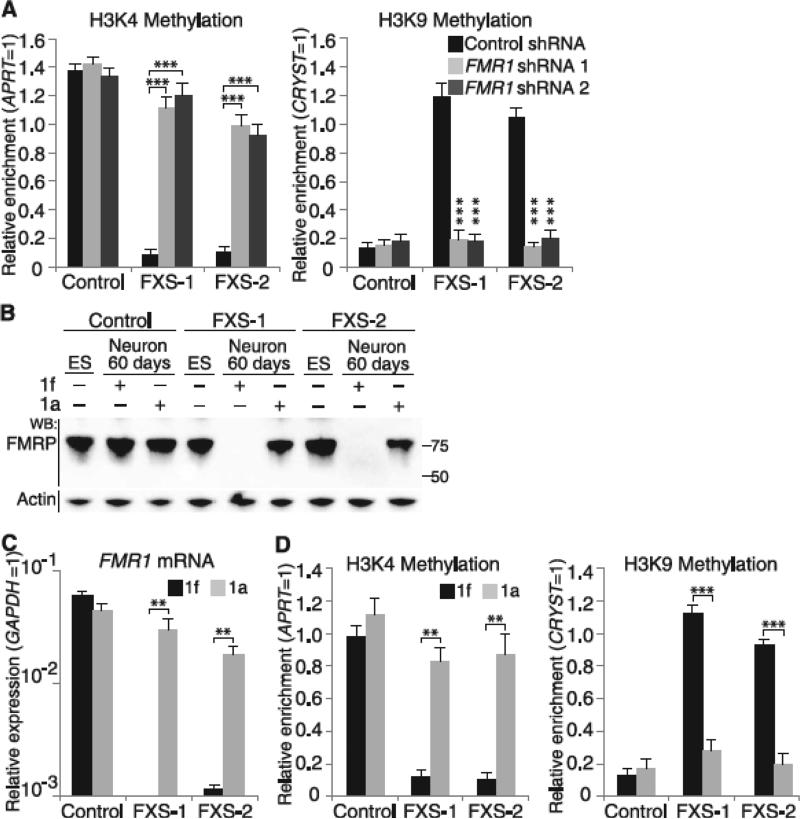
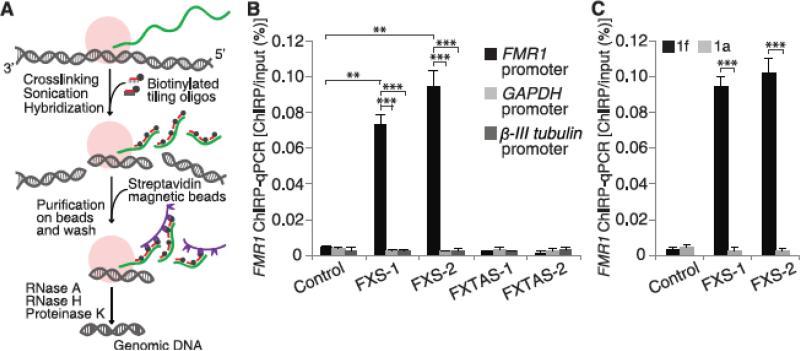
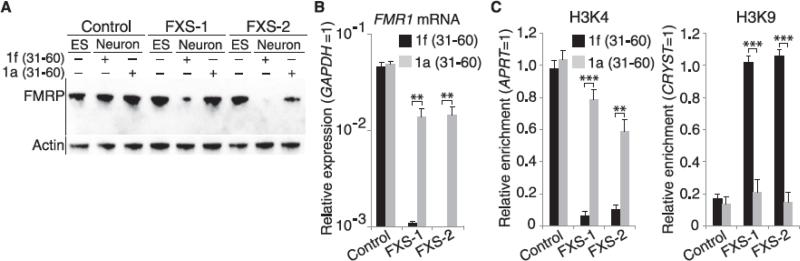
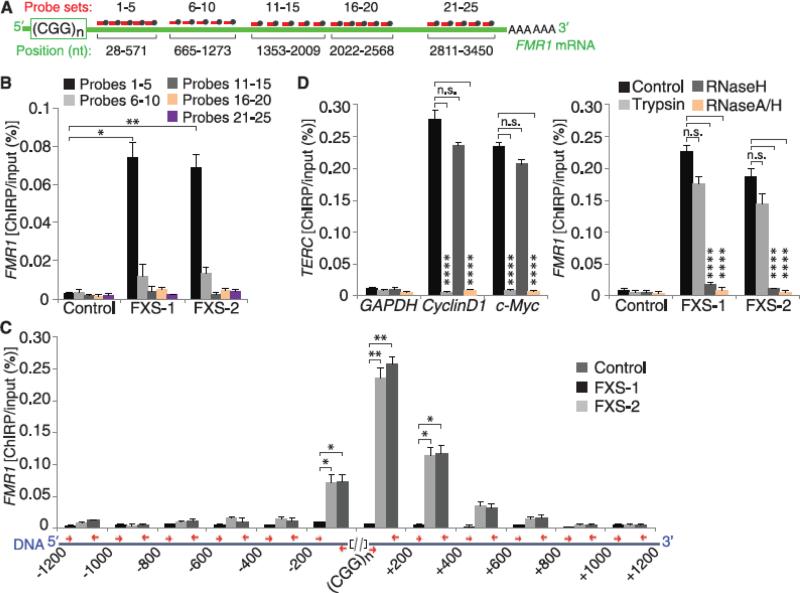
Epigenetic gene silencing is seen in several repeat-expansion diseases. In fragile X syndrome, the most common genetic form of mental retardation, a CGG trinucleotide-repeat expansion adjacent to the fragile X mental retardation 1 (FMR1) gene promoter results in its epigenetic silencing. Here, we show that FMR1 silencing is mediated by the FMR1 mRNA. The FMR1 mRNA contains the transcribed CGG-repeat tract as part of the 5' untranslated region, which hybridizes to the complementary CGG-repeat portion of the FMR1 gene to form an RNA·DNA duplex. Disrupting the interaction of the mRNA with the CGG-repeat portion of the FMR1 gene prevents promoter silencing. Thus, our data link trinucleotide-repeat expansion to a form of RNA-directed gene silencing mediated by direct interactions of the trinucleotide-repeat RNA and DNA.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
